Edit eSignature Form Later
Make the most out of your eSignature workflows with airSlate SignNow
Extensive suite of eSignature tools
Robust integration and API capabilities
Advanced security and compliance
Various collaboration tools
Enjoyable and stress-free signing experience
Extensive support
How To Add Sign in eSignPay

Keep your eSignature workflows on track
Our user reviews speak for themselves









Award-winning eSignature solution




Edit eSignature Form Later. Explore by far the most consumer-warm and friendly knowledge of airSlate SignNow. Control your whole document finalizing and sharing method electronically. Change from hand held, paper-based and erroneous workflows to automated, digital and perfect. It is simple to generate, deliver and indication any files on any product everywhere. Ensure your essential organization cases don't move overboard.
See how to Edit eSignature Form Later. Keep to the simple information to start:
- Make your airSlate SignNow bank account in clicks or log in with your Facebook or Google accounts.
- Enjoy the 30-working day free trial offer or pick a costs plan that's great for you.
- Locate any legitimate format, create on the web fillable types and share them securely.
- Use advanced characteristics to Edit eSignature Form Later.
- Indication, modify putting your signature on order and gather in-particular person signatures ten times more quickly.
- Set automated reminders and get notifications at each and every phase.
Relocating your jobs into airSlate SignNow is simple. What adheres to is a simple process to Edit eSignature Form Later, in addition to ideas to maintain your co-workers and companions for better partnership. Encourage your employees with all the finest tools to remain on top of enterprise functions. Improve output and range your company quicker.
How it works
Rate your experience
-
Best ROI. Our customers achieve an average 7x ROI within the first six months.
-
Scales with your use cases. From SMBs to mid-market, airSlate SignNow delivers results for businesses of all sizes.
-
Intuitive UI and API. Sign and send documents from your apps in minutes.
A smarter way to work: —how to industry sign banking integrate
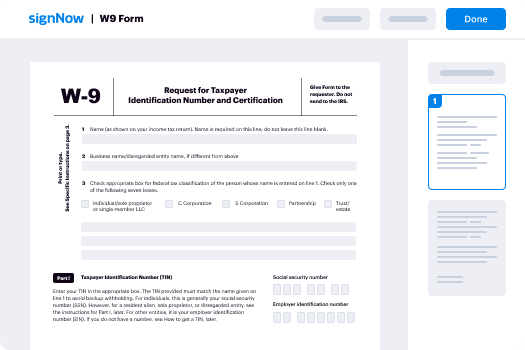
How to sign and fill out a document online
Document management isn't an easy task. The only thing that makes working with documents simple in today's world, is a comprehensive workflow solution. Signing and editing documents, and filling out forms is a simple task for those who utilize eSignature services. Businesses that have found reliable solutions to functionality e signature edit form later don't need to spend their valuable time and effort on routine and monotonous actions.
Use airSlate SignNow and functionality e signature edit form later online hassle-free today:
- Create your airSlate SignNow profile or use your Google account to sign up.
- Upload a document.
- Work on it; sign it, edit it and add fillable fields to it.
- Select Done and export the sample: send it or save it to your device.
As you can see, there is nothing complicated about filling out and signing documents when you have the right tool. Our advanced editor is great for getting forms and contracts exactly how you want/need them. It has a user-friendly interface and full comprehensibility, providing you with total control. Register right now and start enhancing your electronic signature workflows with effective tools to functionality e signature edit form later on the web.
How to sign and complete documents in Google Chrome
Google Chrome can solve more problems than you can even imagine using powerful tools called 'extensions'. There are thousands you can easily add right to your browser called ‘add-ons’ and each has a unique ability to enhance your workflow. For example, functionality e signature edit form later and edit docs with airSlate SignNow.
To add the airSlate SignNow extension for Google Chrome, follow the next steps:
- Go to Chrome Web Store, type in 'airSlate SignNow' and press enter. Then, hit the Add to Chrome button and wait a few seconds while it installs.
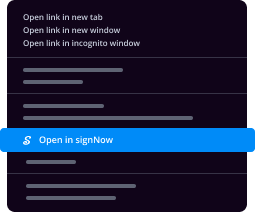
- Find a document that you need to sign, right click it and select airSlate SignNow.
- Edit and sign your document.
- Save your new file to your account, the cloud or your device.
Using this extension, you eliminate wasting time on boring assignments like downloading the file and importing it to a digital signature solution’s library. Everything is easily accessible, so you can quickly and conveniently functionality e signature edit form later.
How to sign docs in Gmail
Gmail is probably the most popular mail service utilized by millions of people all across the world. Most likely, you and your clients also use it for personal and business communication. However, the question on a lot of people’s minds is: how can I functionality e signature edit form later a document that was emailed to me in Gmail? Something amazing has happened that is changing the way business is done. airSlate SignNow and Google have created an impactful add on that lets you functionality e signature edit form later, edit, set signing orders and much more without leaving your inbox.
Boost your workflow with a revolutionary Gmail add on from airSlate SignNow:
- Find the airSlate SignNow extension for Gmail from the Chrome Web Store and install it.
- Go to your inbox and open the email that contains the attachment that needs signing.
- Click the airSlate SignNow icon found in the right-hand toolbar.
- Work on your document; edit it, add fillable fields and even sign it yourself.
- Click Done and email the executed document to the respective parties.
With helpful extensions, manipulations to functionality e signature edit form later various forms are easy. The less time you spend switching browser windows, opening several profiles and scrolling through your internal files looking for a document is more time for you to you for other essential jobs.

How to safely sign documents in a mobile browser
Are you one of the business professionals who’ve decided to go 100% mobile in 2020? If yes, then you really need to make sure you have an effective solution for managing your document workflows from your phone, e.g., functionality e signature edit form later, and edit forms in real time. airSlate SignNow has one of the most exciting tools for mobile users. A web-based application. functionality e signature edit form later instantly from anywhere.
How to securely sign documents in a mobile browser
- Create an airSlate SignNow profile or log in using any web browser on your smartphone or tablet.
- Upload a document from the cloud or internal storage.
- Fill out and sign the sample.
- Tap Done.
- Do anything you need right from your account.
airSlate SignNow takes pride in protecting customer data. Be confident that anything you upload to your account is secured with industry-leading encryption. Automated logging out will protect your account from unauthorized access. functionality e signature edit form later from your mobile phone or your friend’s phone. Security is vital to our success and yours to mobile workflows.

How to eSign a PDF file on an iPhone or iPad
The iPhone and iPad are powerful gadgets that allow you to work not only from the office but from anywhere in the world. For example, you can finalize and sign documents or functionality e signature edit form later directly on your phone or tablet at the office, at home or even on the beach. iOS offers native features like the Markup tool, though it’s limiting and doesn’t have any automation. Though the airSlate SignNow application for Apple is packed with everything you need for upgrading your document workflow. functionality e signature edit form later, fill out and sign forms on your phone in minutes.
How to sign a PDF on an iPhone
- Go to the AppStore, find the airSlate SignNow app and download it.
- Open the application, log in or create a profile.
- Select + to upload a document from your device or import it from the cloud.
- Fill out the sample and create your electronic signature.
- Click Done to finish the editing and signing session.
When you have this application installed, you don't need to upload a file each time you get it for signing. Just open the document on your iPhone, click the Share icon and select the Sign with airSlate SignNow button. Your sample will be opened in the application. functionality e signature edit form later anything. In addition, making use of one service for all your document management needs, everything is faster, smoother and cheaper Download the application today!

How to sign a PDF file on an Android
What’s the number one rule for handling document workflows in 2020? Avoid paper chaos. Get rid of the printers, scanners and bundlers curriers. All of it! Take a new approach and manage, functionality e signature edit form later, and organize your records 100% paperless and 100% mobile. You only need three things; a phone/tablet, internet connection and the airSlate SignNow app for Android. Using the app, create, functionality e signature edit form later and execute documents right from your smartphone or tablet.
How to sign a PDF on an Android
- In the Google Play Market, search for and install the airSlate SignNow application.
- Open the program and log into your account or make one if you don’t have one already.
- Upload a document from the cloud or your device.
- Click on the opened document and start working on it. Edit it, add fillable fields and signature fields.
- Once you’ve finished, click Done and send the document to the other parties involved or download it to the cloud or your device.
airSlate SignNow allows you to sign documents and manage tasks like functionality e signature edit form later with ease. In addition, the security of your information is priority. File encryption and private servers are used for implementing the latest functions in data compliance measures. Get the airSlate SignNow mobile experience and work better.






FAQs
-
What is the process to do e-signature in GST?
E- sign is a new facility provided for GST enrolment. It will enable the taxpayer to sign their GST enrolement application without using DSC. It is however mandatory for some taxpayers to sign GST enrolement applicating using DSC only.Electronically signing of enrolement application using DSC is mandatory for:CompaniesForeign companiesLi mited liability partnership (LLP)Foreign limited liability partnership (FLLP’s)Only the taxpayers other than mentioned above will be able to file their enrolment application without using DSC. If you are an individual , HUF and partnership then you can file the enrolment application without DSC. You can sign the enrolment application electronically using the E- sign. It will result in cost saving as you will not have pay for the DSC. It will be easier for you as other procedure like registering DSC will also not be required.What is E- sign? How does it work?E- sign stands for electronic signature. It is an online electronic signature service that allows an Aadhaar holder to digitally sign a document. If the taxpayer opts to electronically sign the enrolement application or any other document at the GST common portal using the e- sign services.Following steps will be required to use E- signThe GST common portal prompts the taxpayer to enter the Aadhaar number of the authorized signatory.After validating the Aadhar number , the GST common portal sends a request to UIDAI system to send an OTPUIDAI system sends an OTP to e- mail address and mobile number registered against Aadhar number.the GST system prompts the taxpayer to enter the OTP.The taxpayer enters the OTP and submits the Enrollment Application or the document. The E- signing process is completed.This facility is free of cost and easy to use.It is mandatory to file your application with digital sign. This sign can be via DSC or E-sign. You will not be able to file your enrolement application without signature.
-
How do I create a web design contract?
First of all, good for you for having the mental clarity to want to use contracts for web design. You believe it or not, are far ahead of most “developers” just by virtue of being able to conceive of using contracts. You’ll be ahead of most “wantrepreneurs” and “startup founders” just by thinking of using contracts, so hats off to you young man.You can find sample contracts very easily, just by Googling the free-to-use “contract killer” template. Just Google “contract killer web design contract.” Another way to find even more sample contracts is to take a few “how to be a web developer” courses on Udemy and I think you’d probably find more by Googling “sample web design contract,” going to the Freelancer’s Union website, going to signNow and looking at theirs as well.I’m not a lawyer, but I don’t have to be one (nor do you) in order to use or edit contracts. I started out as a web developer not using contracts simply because nobody else I spoke with knew anything and it was a very intimidating topic; and like so many other things of value in life, if you want it done correctly sometimes you just have to get off your butt and do some hard work. I had more than my fair share of being taken advantage of by non-profits (I used to be in favor of volunteering, now I will never do it again) who once they had pro bono (free basically) work, had “revisions” without end - taking professional design and wanting to turn it into a coloring book for a bi-polar paranoid schizophrenic. I had “agencies” asking me to build sites for them “on spec,” (for free), playing mind games with “more work later.” “Agencies” with $200 budgets, and so on and on it went. Until I saw what real agencies charged for crap work with no SEO or thought. Now I always contracts, and I use electronic signatures, and store contracts for years on Dropbox in case they “forget” something a year or two out.I keep a folder of different contracts, RFP responses, generic “sorry your budget is so low” letters, and “hey, we’d love to work with you” letters and suggest you do the same. You can use Gravity Forms to set up or send form contracts with IP tracking and e-signature or use fill-in PDF forms, or even old-fashioned paper contracts, but contracts are a necessity especially in a profession as under-valued and as misunderstood as ours.Good luck and if you need any future help, I’d be happy to pitch in.
-
What is the best email hosting besides Google?
Disclaimer: we do not work at Zoho. We are Freelensia, an online platform that connects freelance interpreters and companies looking for short-term interpretation for meetings, seminars and exhibitions. Our moto is interpreters anywhere, anytime, for any language. If you are interested, please visit us at https://www.freelensia.com.We are a previous user of Google’s corporate mail and currently are using Zoho Mail. Please see the screenshot below:The interface is very much like Gmail. We had some trouble visually in the beginning because it is so overwhelming (the view seems more cluttered than Gmail). But we later appreciate very much because it is a lot more powerful. For example, you can tag people in your organization in a specific mail instead of forwarding the e-mail.A big plus is that you will be able to integrate with a entire suite of Zoho products: forms, campaigns, CRM etc. Some of these products do not exist for Google or is a lot better.A drawback of Zoho Mail is that you do not have all the cool labs of Google. Setting up signatures is a bit difficult. Canned responses do not exist. There may be some font issues if you are writing e-mails in non-alphabetic languages.The coolest thing is that their basic plan is free forever up to 15 e-mail addresses. I don't think there are many e-mail services out there that offer this level of flexibility for early startups.If you are interested, please use the link below to sign up. We will both receive 5 more free e-mail address slots from this invite. It is completely and forever free as long as you are below the limit for number of users. No trial, no credit card input, etc.EDIT: The limit is now 25 users + 5 free users from our referral linkLet me know if you have issues setting up!https://zm4.bz/7M01kkXa
-
How can I register for SSC CGL/SSC CHSL 2017 exam?
All the interested candidates who are looking for all the important details regarding SSC CGL 2017, they can now check out the same from the official website, however the exact dates for several activities regarding SSC CGL 2017 is not out yet. SSC is itself a huge name. Each year, it recruits various vacancies for the several posts. As per the sources, the SSC CGL 2017 examinations may conduct in the month of Aug – Sept. All the interested candidates are advised to have a look on the given details so that they can know, whether they are eligible or not to apply for this examination.Organization’s Name: Staff Selection Commission (SSC)Examination’s Name: Combined Graduate Level Examination(CGLE)The Application form will be available from: February 2017Mode of Apply: Online Mode and Offline ModeMode of Exam: Computer Based Test (CBT)Application form will be available from -February 2017Last date to apply-March 2017Admit card will be available for SSC CGL Tier-1-April 2017SSC CGL Tier-1 Exam-May 2017Result declaration for Tier-June 2017Admit card will be available for SSC CGL Tier-2-July 2017SSC CGL Tier-2 Exam-August 2017Result declaration for CGL Tier-2-September 2017SSC CGL Tier-3 Exam-October 2017Interview/Skill Test/Computer Skill Test-November 2017Eligibility CriteriaEducation: Candidate must have done graduation in any stream from a recognised university.Age Limit – Candidate’s age must be between 18 to 27 years old to be eligible for this examination.How to ApplyFirst of all, all the candidates are requested to visit the official website that is Once you are on the home page, then you will have to open the SSC CGL Apply online link (which will be active after release of official notice for SSC CGL 2017 Examination).Read all the instructions carefully and then fill the application form. Make sure all the details are filled accurately and completely. Once the details are filled in, then you will be asked to upload the scanned photograph and signature in prescribed format. Check all the details, if they are mentioned correctly. Lastly, just submit the form. Don’t forget to take the print out of the confirmation page.
-
Is Evolution founded on scientific proof, or is it a godless worldview founded on supposition?
Science does not operate on a 'proof' model, it is a falsification model where the hypothesis that is best supported by mountains of evidence and not falsified are promoted to Scientific Theory status.Here are just a tiny few of the many tens of thousands of studies that support Evolutionary theory - each such paper contains a record of the scientific methodology applied and the scientific evidence produced so it can be challenged or reproduced by other researchers.Theobald, D. L. 2010. A formal test of the theory of universal common ancestry. Nature 465:219-223Adl, S., Leander, B.S., Simpson, A.G.B., Archibald, J.M., Anderson, O.R., Bass, D., Bowser, S.S., Brugerolle, G., Farmer, M.A., Karpov, S., Kolisko, M., Lane, C.E., Lodge, D.J., Mann, D.G., Meisterfeld, R., Mendoza, L., Moestrup, Ø., Mozley-Standridge, S.E., Smirnov, A.V., and Spiegel, F. (2007) Diversity, nomenclature, and taxonomy of protists. Syst. Biol., 56, 684-689.Adl, S.M., Simpson, A.G., Farmer, M.A., Andersen, R.A., Anderson, O.R., Barta, J.R., Bowser, S.S., Brugerolle, G., Fensome, R.A., Fredericq, S., James, T.Y., Karpov, S., Kugrens, P., Krug, J., Lane, C.E., Lewis, L.A., Lodge, J., Lynn, D.H., Mann, D.G., McCourt, R.M., Mendoza, L., Moestrup, O., Mozley-Standridge, S.E., Nerad, T.A., Shearer, C.A., Smirnov, A.V., Spiegel, F.W. and Taylor, M.F. (2005) The new higher level classification of eukaryotes with emphasis on the taxonomy of protists. J. Eukaryot. Microbiol., 52, 399-451.Adoutte, A., G. Balavoine, N. Lartillot, O. Lespinet, B. Prud'homme, and R. de Rosa. 2000. The new animal phylogeny: Reliability and implications. Proceedings of the National Academy of Sciences (USA) 97:4453-4456.Ahmad, S., A. Selvapandiyan, and R. K. Bhatnagar. 1999. A protein-based phylogenetic tree for Gram-positive bacteria derived from hrcA, a unique heat-shock regulatory gene. International Journal of Systematic Bacteriology 49:1387-1394.Anderson, C. L. 1998. Phylogenetic relationships of the Myxozoa. Pages 341-350 in Evolutionary Relationships among Protozoa (G.H. Coombs, K. Vickerman, M.A. Sleigh, and A. Warren, eds.) Chapman & Hall, London.Anderson, C. L., E. U. Canning, and B. Okamura. 1998. A triploblast origin for Myxozoa? Nature 392:346-347.Andersson, S. G. E., A. Zomorodipour, J. O. Andersson, T. Sicheritz-Ponten, U. C. M. Alsmark, R. M. Podowski, A. K. Naslund, A. S. Eriksson, H. H. Winkler, and C. G. Kurland. 1998. The genome sequence of Rickettsia prowazekii and the origin of mitochondria. Nature 396:133-140.Andersson, S.G. and Kurland, C.G. (1999) Origins of mitochondria and hydrogenosomes. Curr. Opin. Microbiol., 2, 535-541.Aravind, L., R. L. Tatusov, Y. I. Wolf, D. R. Walker, and E. V. Koonin. 1998. Evidence for massive gene exchange between archaeal and bacterial hyperthermophiles. Trends in Genetics 14:442-444.Archibald, J.M. (2005) Jumping genes and shrinking genomes.probing the evolution of eukaryotic photosynthesis with genomics. IUBMB Life, 57, 539-547.Archibald, J.M., Longet, D., Pawlowski, J. and Keeling, P.J. (2002) A novel polyubiquitin structure in Cercozoa and Foraminifera: evidence for a new eukaryotic supergroup. Mol. Biol. Evol., 20, 62-66.Arisue, N., Hasegawa, M., and Hashimoto, T. (2005) Root of the Eukaryota tree as inferred from combined maximum likelihood analyses of multiple molecular sequence data. Molecular Biology and Evolution, 22(3), 409-420.Ayala, F. J., A. Rzhetsky, and F. J. Ayala. 1998. Origin of the metazoan phyla: Molecular clocks confirm paleontological estimates. PProceedings of the National Academy of Sciences (USA) 95:606-611.Baguñà, J., P. Martinez, J. Paps, and M. Riutort. 2008. Back in time: a new systematic proposal for the Bilateria. Philosophical Transactions of the Royal Society Series B 363(1496):1481-1491Baldauf, S. L. (1999) A search for the origins of animals and fungi: Comparing and combining molecular data. American Naturalist, 154(suppl.), S178-S188.Baldauf, S. L., J. D. Palmer, and W. F. Doolittle. 1996. The root of the universal tree and the origin of eukaryotes based on elongation factor phylogeny. Proceedings of the National Academy of Sciences of the United States of America 93:7749-7754.Baldauf, S.L. and Doolittle, W.F. (1997) Origin and evolution of the slime molds (Mycetozoa). Proceedings of the National Academy of Sciences (USA), 94, 12007-12012.Baldauf, S.L. and Palmer, J.D. (1993) Animals and fungi are each other's closest relatives: congruent evidence from multiple proteins. Proc. Natl. Acad. Sci. USA, 90, 11558-11562.Baldauf, S.L., Roger, A.J., Wenk-Siefert, I. and Doolittle, W.F. (2000) A kingdom-level phylogeny of eukaryotes based on combined protein data. Science, 290, 972-977.Balows, A., H.G. Träper, M. Dworkin, W. Harder, and K.-H. Schleifer (eds.). 1992. The Prokaryotes. A Handbook on the Biology of Bacteria: Ecophysiology, Isolation, Identification, Applications. Second edition, Volumes I-IV. Springer Verlag, New York.Bapteste, E., Brinkmann, H., Lee, J., Moore, D., Sensen, C., Gordon, P., Durufle, L., Gaasterland, T., Lopez, P., Muller, M. and Philippe, H. (2002) The analysis of 100 genes supports the grouping of three highly divergent amoebae: Dictyostellium, Entamoeba, and Mastigamoeba. Proc. Natl. Acad. Sci. U S A, 99, 1414-1419.Barns, S. M., C. F. Delwiche, J. D. Palmer, and N. R. Pace. 1996. Perspectives on archaeal diversity, thermophily and monophyly from environmental rRNA sequences. Proceedings of The National Academy of Sciences (U.S.A.) 93:9188-9193.Barns, S. M., R. E. Fundyga, M. W. Jeffries and N. R. Pace. 1994. Remarkable archaeal diversity detected in a Yellowstone National Park hot spring environment. Proceedings of the National Academy of Sciences of the United States of America 91(5): 1609-1613.Bass, D., Moreira, D., Lopez-Garcia, P., Polet, S., Chao, E.E., von der Heyden, S., Pawlowski, J. and Cavalier-Smith, T. (2005) Polyubiquitin insertions and the phylogeny of Cercozoa and Rhizaria. Protist, 156, 149-161.Battistuzzi, F. U. and A. B. Hedges. 2009. A major clade of prokaryotes with ancient adaptations to life on land. Molecular Biology and Evolution 26(2):335-343; doi:10.1093/molbev/msn247Battistuzzi, F. U., A. Feijao, and A. B. Hedges. 2004. A genomic timescale of prokaryote evolution: insights into the origin of methanogenesis, phototrophy, and the colonization of land. BMC Evolutionary Biology 4:44-.Becerra, A., L. Delaye, S. Islas, and A. Lazcano. 2007. The very early stages of biological evolution and the nature of the last common ancestor of the three major cell domains. Annual Review of Ecology, Evolution, and Systematics 38:361-379.Benachenhou, L. N., P. Forterre and B. Labedan. 1993. Evolution of glutamate dehydrogenase genes: Evidence for two paralogous protein families and unusual branching patterns of the archaebacteria in the universal tree of life. Journal Of Molecular Evolution 36(4): 335-346.Benachenhou, L. N., P. Forterre and B. Labedan. 1993. Evolution of glutamate dehydrogenase genes: Evidence for two paralogous protein families and unusual branching patterns of the archaebacteria in the universal tree of life. Journal Of Molecular Evolution 36:335-346.Bern, M. and D. Goldberg. 2005. Automatic selection of representative proteins for bacterial phylogeny. BMC Evolutionary Biology 5:34-.Berney, C. and Pawlowski, J. (2006) A molecular time-scale for eukaryote evolution recalibrated with the continuous microfossil record. Proceedings of the Royal Society Series B, 273(1596), 1867-1872.Boone, D. R., R.W. Castenholz, and G.M. Garrity. 2001. Bergey's Manual of Systematic Bacteriology. Springer, New York.Borchiellini C., Boury-Esnault, N., Vacelet, J., and Le Parco, Y. (1998) Phylogenetic analysis of the Hsp70 sequences reveals the monophyly of metazoa and specific phylogenetic relationships between animals and fungi. Molecular Biology and Evolution, 15, 647-655.Borchiellini, C., M. Manuel, E. Alivon, N. Boury-Esnault, J. Vacelet, and Y. Le Parco. 2001. Sponge paraphyly and the origin of Metazoa. Journal of Evolutionary Biology 14:171-179.Briggs, D. E. G., D. H. Erwin, and F. J. Collier. 1994. The Fossils of the Burgess Shale. Smithsonian Institution Press, Washingthon, D.C.Brinkmann, H. and H. Phillippe. 1999. Archaea sister group of bacteria? Indications from Tree Reconstruction Artifacts from ancient Phylogenies. Molecular Biology and Evolution 16:817-825.Brochier, C., E. Bapteste, D. Moreira, and H. Philippe. 2002. Eubacterial phylogeny based on translational apparatus proteins.Brocks, J. J., G. A. Logan, R. Buick, and R. E. Summons. 1999. Archean molecular fossils and the early rise of eukaryotes. Science 285:1033-1036.Brown, J. R. , C. J. Douady, M. J. Italia, W. E. Marshall, and M. J. Stanhope. 2001. Universal trees based on large combined protein sequence data sets. Nature Genetics 28:281-285.Brown, J. R. 2001. Genomic and phylogenetic perspectives on the evolution of prokaryotes. Systematic Biology 50:497-512.Brown, J. R. and W. F. Doolittle. 1995. Root of the universal tree of life based on ancient aminoacyl-tRNA synthetase gene duplications. Proceedings of the National Academy of Sciences of the United States of America 92:2441-2445.Brown, J. R. and W. F. Doolittle. 1997. Archaea and the prokaryote-to-eukaryote transition. Microbiology and Molecular Biology Reviews 61:456-502.Brusca, R. C. and G. J. Brusca. 2002. Invertebrates. Second Edition. Sinauer Associates, Inc., Sunderland, Massachusetts.Budd, G. E. 2008. The earliest fossil record of the animals and its significance. Philosophical Transactions of the Royal Society Series B 363(1496):1425-1434.Budin, K. and Philippe, H. (1998) New insights into the phylogeny of eukaryotes based on Ciliate Hsp70 sequences. Molecular Biology and Evolution, 15, 943-956.Burki, F. and Pawlowski, J. (2006) Monophyly of Rhizaria and multigene phylogeny of unicellular bikonts. Molecular Biology and Evolution, 23(10), 1922-1930.Burki, F., Shalchian-Tabrizi, K. and Pawlowski, J. (2008) Phylogenomics reveals a new 'megagroup' including most photosynthetic eukaryotes. Biol. Lett., 4(4), 366-369.Burki, F., Shalchian-Tabrizi, K., Minge, M., Skjaeveland, A., Nikolaev, S.I., Jakobsen, K.S. and Pawlowski, J. (2007) Phylogenomics reshuffles the eukaryotic supergroups. PLoS ONE, 2, e790.Bustard, K. and R. S. Gupta. 1997. The sequences of heat shock protein 40 (DnaJ) homologs provide evidence for a close evolutionary relationship between the Deinococcus-Thermus group and cyanobacteria. Journal of Molecular Evolution 45:193-205.Caetano-Anolles, G. 2002. Evolved RNA secondary structure and the rooting of the universal tree of life. Journal of Molecular Evolution 54: 333-345.Calisher, C. H., M. C. Horzinek, M. A. Mayo, H. W. Ackermann, and J. Maniloff. 1995. Sequence analyses and a unifying system of virus taxonomy - consensus via consent. Archives of Virology 140 (11):2093-2099.Cammarano, P., P. Palm, R. Creti, E. Ceccarelli, A. M. Sanangelantoni, and O. Tiboni. 1992. Early evolutionary relationships among known life forms inferred from elongation factor EF-2/EF-G sequences: Phylogenetic coherence and structure of the Archaeal domain. Journal Of Molecular Evolution 34:396-405.Cammarano, P., R. Creti, A. M. Sanangelantoni, and P. Palm. 1999. The archaean monophyly issue: a phylogeny of translational elongation factor G(2) sequences inferred from an optimized selection of alignment positions. Journal Of Molecular Evolution 49:524-537.Canning, E.U. (1998) Evolutionary relationships of Microsporidia. Pages 77-90 in Evolutionary Relationships among Protozoa (G. H. Coombs, K. Vickerman, M .A. Sleigh, and A. Warren, eds.) Chapman & Hall, London.Carroll, S. B., J. K. Grenier, and S. D. Weatherbee. 2001. From DNA to Diversity. Molecular Genetics and the Evolution of Animal Design. Blackwell Science, Malden, Massachusetts.Castro, H. F., N. H. Williams, and A. Ogram. 2000. Phylogeny of sulfate-reducing bacteria. FEMS Microbiology Ecology 31:1-9.Cavalier-Smith, T. (1987) The origin of fungi and pseudofungi. In Rayner, A.D.M., Brasier, C.M. and Moore, D. (eds.), Evolutionary biology of the fungi. Cambridge University Press, Cambridge, pp. 339-353.Cavalier-Smith, T. (1993) Kingdom Protozoa and its 18 phyla. Microbiol. Rev., 57, 953-94.Cavalier-Smith, T. (1998) A revised six-kingdom system of life. Biol. Rev. Camb. Philos. Soc., 73, 203-266.Cavalier-Smith, T. (1999) Principles of protein and lipid targeting in secondary symbiogenesis: euglenoid, dinoflagellate, and sporozoan plastid origins and the eukaryote family tree. J. Eukaryot. Microbiol., 46, 347-366.Cavalier-Smith, T. (2002) The phagotrophic origin of eukaryotes and phylogenetic classification of Protozoa. Int. J. Sys. Evol. Microbiol., 52, 297-354.Cavalier-Smith, T. (2003) Protist phylogeny and the high-level classification of Protozoa. Eur. J. Protistol., 39, 338-348.Cavalier-Smith, T. (2004) Chromalveolate diversity and cell megaevolution: interplay of membranes, genomes and cytoskeleton. In Hirt, R.P. and Horner, D. (eds.), Organelles, Genomes and Eukaryotic Evolution. Taylor and Francis, London, pp. 71-103.Cavalier-Smith, T. 2002. The neomuran origin of archaebacteria, the negibacterial root of the universal tree and bacterial megaclassification. International Journal of Systematic and Evolutionay Microbiology 52:7-76.Cavalier-Smith, T. and Chao, E.E. (1995) The opalozoan Apusomonas is related to the common ancestor of animals, fungi and choanoflagellates. Proceedings of the Royal Society of London Series B, 261, 1-6.Cavalier-Smith, T. and Chao, E.E. (2003) Phylogeny and classification of phylum Cercozoa (Protozoa). Protist, 154, 341-358.Cavalier-Smith, T., M. T. E. P. Allsopp, E. E. Chao, N. Boury-Esnault, and J. Vacelet. 1996. Sponge phylogeny, animal monophyly, and the origin of the nervous system: 18S rRNA evidence. Canadian Journal of Zoology 74:2031-2045.Chen, J.-Y., P. Oliveri, C.-W. Li, G.-Q. Zhou, F. Gao, J. W. Hagadorn, K. J. Peterson, and E. H. Davidson. 2000. Precambrian animal diversity: Putative phosphatized embryos from the Doushantuo Formation of China. Proceedings of the National Academy of Sciences (U.S.A.) 97:4457-4462.Ciccarelli, F. D., T. Doerks, C. von Mering, C. J. Creevey, B. Snel, and P. Bork. 2006. Toward automatic reconstruction of a highly resolved tree of life. Science 311(5765):1283-1287.Clark C.G. and Roger, A.J. (1995) Direct evidence for secondary loss of mitochondria in Entamoeba histolytica. Proceedings of the National Academy of Sciences (USA), 92, 6518-6521.Coenye, T. and P. Vandamme. 2004. A genomic perspective on the relationship between the Aquificales and the epsilon-Proteobacteria. Syst Appl. Microbiol. 27:313-322.Collins, A. G. 1998. Evaluating multiple alternative hypotheses for the origin of Bilateria: An analysis of 18S rRNA molecular evidence. Proceedings of the National Academy of Sciences (U.S.A.) 95:15458-15463.Collins, A. G. and J. W. Valentine. 2001. Defining phyla: evolutionary pathways to metazoan body plans. Evolution & Development 3:432-442.Conway Morris, S. 1993. The fossil record and the early evolution of the Metazoa. Nature 361:219-225.Conway Morris, S. 1998. The Crucible of Creation: The Burgess Shale and the Rise of Animals. Oxford University Press, Oxford, UK.Copeland, H.F. (1956) The Classification of Lower Organisms. Pacific Books, Palo Alto, California.Copley, R. R. 2008. The animal in the genome: comparative genomics and evolution. Philosophical Transactions of the Royal Society Series B 363(1496):1453-1461.Creti, R., E. Ceccarelli, M. Bocchetta, A. M. Sanangelantoni, O. Tiboni, P. Palm and P. Cammarano. 1994. Evolution of translational elongation factor (EF) sequences: Reliability of global phylogenies inferred from EF-1-alpha(Tu) and EF-2(G) proteins. Proceedings of the National Academy of Sciences of the United States of America 91:3255-3259.Daubin, V., M. Gouy, and G. Perriere. 2002. A phylogenomic approach to bacterial phylogeny: Evidence of a core of genes sharing a common history. Genome Research 2(7):1080-1090.DeLong E. F. and N. R. Pace. 2001. Environmental diversity of Bacteria and Archaea. Systematic Biology 50: 470-478.DeLong E. F. and N. R. Pace. 2001. Environmental diversity of Bacteria and Archaea. Systematic Biology 50:470-478.Deeds, E. J., H. Hennessey, and E. I. Shakhnovich. 2005. Prokaryotic phylogenies inferred from protein structural domains. Gen. Res. 15:393-402.Dellaporta, S. L., A. Xu, S. Sagasser, W. Jakob, M. A. Moreno, L. W. Buss, and B. Schierwater. 2006. Mitochondrial genome of Trichoplax adhaerens supports placozoa as the basal lower metazoan phylum. Proceedings of the National Academy of Sciences (U.S.A.) 103(23):8751-8756.Delong, E. F. 1992. Archaea in coastal marine environments. Proceedings of The National Academy of Sciences (U.S.A.) 89: 5685-5689.Delwiche, C.F. (1999) Tracing the thread of plastid diversity through the tapistry of life. American Naturalist, 154 (suppl.), S164-S177.Des Marais, D. J. 1999. Astrobiology: Exploring the origins, evolution, and distribution of life in the universe. Annual Review of Ecology and Systematics 30:397-420.Dewel, R. A. 2000. Colonial origin for Eumetazoa: Major morphological transitions and the origin of Bilaterian complexity. Journal of Morphology 243:35-74.Doolittle, W. F. 1998. You are what you eat: a gene transfer ratchet could account for bacterial genes in eukaryotic nuclear genomes. Trends in Genetics 14:307-311.Doolittle, W. F. 1999. Lateral genomics. Trends in Biochemical Sciences 24: M5-M8.Doolittle, W. F. 1999. Phylogenetic classification and the universal tree. Science 284:2124-2128.Doolittle, W. F. 2000. Uprooting the tree of life. Scientific American 282:90-95.Doolittle, W. F. and J. M. Logsdon. 1998. Archaeal genomics: Do archaea have a mixed heritage? Current Biology 8: (6) R209-R211.Doolittle, W. F. and J. R. Brown. 1994. Tempo, mode, the progenote, and the universal root. Proceedings of the National Academy of Sciences of the United States of America 91:6721-6728.Douzery, E.J.P., Snell, E.A., Bapteste, E., Delsuc, F., and Philippe, H. (2004) The timing of eukaryotic evolution: Does a relaxed molecular clock reconcile proteins and fossils? Proceedings of the National Academy of Sciences (USA), 101(43), 15386-15391.Dunn, C. W., A Hejnol, D. Q. Matus, K. Pang, W. E. Browne, S. A. Smith, E. Seaver, G. W. Rouse, M. Obst, G. D. Edgecombe, M. V. Sørensen, S. H. D. Haddock, A. Schmidt-Rhaesa, A. Okusu, R. M. Kristensen, W. C. Wheeler, M. Q. Martindale, and G. Giribet. 2008. Broad phylogenomic sampling improves resolution of the animal tree of life. Nature. doi:10.1038/nature06614Edlind, T.D. (1998) Phylogenetics of protozoan tubulin with reference to the amitochondriate eukaryotes. Pages 91-108 in Evolutionary Relationships Among Protozoa (Coombs, G.H., Vickerman, K., Sleigh, M.A. and Warren, A., eds.) Chapman & Hall, London.Edlind, T.D., Li, J., Visvesvara, G.S., Vodkin, M.H., McLaughlin, G.L., and Katiyar, S.K. (1996) Phylogenetic analysis of beta-tubulin sequences from amitochondrial protozoa. Molecular Phylogenetics and Evolution, 5, 359-367.Eernisse, D. J. and K. J. Peterson. 2004. The history of animals. Pages 197-208 in Assembling the Tree of Life, J. Cracraft and M. J. Donoghue, eds. Oxford University Press, New York.Eisen, J. A. 1995. The RecA protein as a model molecule for molecular systematic studies of bacteria: Comparison of trees of RecAs and 16S rRNAs from the same species. Journal of Molecular Evolution 41:1105-1123. Molecular Biology and Evolution 21(9):1643-1660.Embley, T. M., M. van der Giezen, D. S. Horner, P. L. Dyal, S. Bell, and P. G. Foster. 2003. Hydrogenosomes, mitochondria and early eukaryotic evolution. International Union of Biochemistry and Molecular Biology: Life 55(7):387-395.Embley, T.M. (2006) Multiple secondary origins of the anaerobic lifestyle in eukaryotes. Philos. Trans. R. Soc. Lond. B Biol. Sci., 361, 1055-1067.Embley, T.M. and Hirt, R.P. (1998) Early branching eukaryotes? Curr. Opinion Gen. Dev., 8, 624-629.Emelyanov, V. V. and B. V. Sinitsyn. 1999. A groE-based phylogenetic analysis shows very close evolutionary relationship between mitochondria and Rickettsia. Russian Journal of Genetics 35:618-627.Ender, A. and B. Schierwater. 2003. Placozoa are not derived cnidarians: Evidence from molecular morphology. Molecular Biology and Evolution 20(1):130-134.Erwin, D. H. 1993. The origin of metazoan development: A palaeobiological perspective. Biological Journal of the Linnean Society 50: 255-274.Esser, C., N. Ahmadinejad, C. Wiegand, C. Rotte, F. Sebastiani, G. Gelius-Dietrich, K. Henze, E. Kretschmann, E. Richly, D. Leister, D. Bryant, M. A. Steel, P. J. Lockhart, D. Penny and W. Martin. 2004. A genome phylogeny for mitochondria among alpha-proteobacteria and a predominantly eubacterial ancestry of yeast nuclear genes.Fast, N.M., Kissinger, J.C., Roos, D.S. and Keeling, P.J. (2001) Nuclear-encoded, plastid-targeted genes suggest a single common origin for apicomplexan and dinoflagellate plastids. Mol. Biol. Evol., 18, 418-426.Fast, N.M., Logsdon, J.M., and Doolittle, W.F. (1999) Phylogenetic analysis of the TATA box binding protein (TBP) gene from Nosema locustae: evidence for a microsporidia-fungi relationship and spliceosomal intron loss. Molecular Biology and Evolution, 16, 1415-1419.Fauquet, C. M., M. A. Mayo, J. Maniloff, U. Desselberger, and L. A. Ball (Eds.). 2005. Virus Taxonomy. Elsevier, San Diego.Feng, D.-F., G. Cho, and R.F. Doolittle. 1997. Determining divergence times with a protein clock: Update and reevaluation. Proceedings of the National Academy of Sciences of the United States of America 94:13028-13033.Ferrier, D. E. K. and P. W. H. Holland. 2001. Ancient origin of the Hox gene cluster. Nature Reviews Genetics 2:33-38.Fields, B. N., D. M. Knipe, and P. M. Howley (eds.) 1996. Fields Virology, 3rd ed. Lippincott-Raven Publishers, Philadelphia, Pa.Finnerty, J. R., K. Pang, P. Burton, D. Paulson, and M. Q. Martindale. 2004. Origins of bilateral symmetry; Hox and dpp expression in a sea anemone. Science 304:1335-37.Forterre, P. 2001. Genomics and early cellular evolution. The origin of the DNA world. Comptes Rendus de l'Academie des Sciences Serie III-Sciences de la Vie 324:1067-1076.Forterre, P. and H. Philippe. 1999. Where is the root or the universal tree of life? BioEssays 21:871-879.Forterre, P., Benachenhou-Lahfa, N., Confalonieri, F., Duguet, M., Elie, C., and Labedan, B. (1992) The nature of the last universal ancestor and the root of the tree of life, still open questions. Biosystems, 28, 15-32.Fox, G. E., E. Stackebrandt, R. B. Hespell, J. Gibson, J. Maniloff, T. A. Dyer, R. S. Wolfe, W. E. Balch, R. S. Tanner, L. J. Magrum, L. B. Zablen, R. Blakemore, R. Gupta, L. Bonen, B. J. Lewis, D. A. Stahl, K. R. Luehrsen, K. N. Chen, and C. R. Woese. 1980. The phylogeny of prokaryotes. Science 209:457-463.Garrity, G. M., J. A. Bell, and T. G. Lilburn. 2004. Taxonomic Outline of the Prokaryotes. Bergey's Manual of Systematic Bacteriology, Second Edition. Release 5.0.Germot, A., Philippe, H., and Le Guyader, H. (1997) Evidence for loss of mitochondria in Microsporidia from a mitochondrial-type HSP70 in Nosema locustae. Molecular and Biochemical Parasitology, 87, 159-168.Gibbs, A. J. 2000. Virus nomenclature descending into chaos. Archives of Virology 145:1505-1507.Giribet, G. 2002. Current advances in the phylogenetic reconstruction of metazoan evolution. A new paradigm for the Cambrian explosion? Molecular Phylogenetics and Evolution 24:345-357.Giribet, G., C. W. Dunn, G. D. Edgecombe, and G. W. Rouse. 2007. A modern look at the Animal Tree of Life. Pages 61-79 in: Zhang, Z.-Q. & Shear, W.A., eds. Linnaeus Tercentenary: Progress in Invertebrate Taxonomy. Zootaxa 1668:1-766.Gogarten, J. P. and L. Taiz. 1992. Evolution of proton pumping ATPases: Rooting the tree of life. Photosynthesis Research 33:137-146.Gogarten, J. P., E. Hilario, and L. Olendzenski. 1996. Gene duplications and horizontal gene transfer during early evolution. Pages 267-292 in Evolution of Microbial Life (D. McL. Roberts, P. Sharp, G. Alderson, and M. Collins, eds.) Symposium 54. Society for General Microbiology. Cambridge University Press, Cambridge.Gogarten, J.P. (2003) Gene transfer: Gene swapping craze signNowes eukaryotes. Curr Biol., 13, R53.54.Gogarten, J.P., Kiblak, H., Dittrich, P., Taiz, L., Bowman, E.J., Bowman, B.J., Manolson, N.F., Poole, R.J., Date, T., Oshima, T., Konishi, J., Denda, K. and Yoshida, M. (1989) Evolution of the vacuolar H+-ATPase: inplications for the origin of eukaryotes. Proc. Natl. Acad. Sci. USA, 86, 6661-6665.Golding, G.B. and Gupta, R. S. (1995) Protein-based phylogenies support a chimeric origin for the eukaryotic genome. Molecular Biology and Evolution, 12, 1-6.Golding, G.B. and R.S. Gupta. 1995. Protein-based phylogenies support a chimeric origin for the eukaryotic genome. Molecular Biology and Evolution 12:1-6.Gould, S. J. 1989. Wonderful Life: The Burgess Shale and the Nature of History. Norton, New York.Gould, S.B., Waller, R.F. and McFadden, G.I. (2008) Plastid evolution. Annu. Rev. Plant. Biol., 59, 491-517.Gouy, M. and W.-H. Li. 1989. Phylogenetic analysis based on rRNA sequences supports the archaebacterial rather than the eocyte tree. Nature 339:145-147.Gouy, M. and W.-H. Li. 1990. Archaebacterial or eocyte tree? Nature 343:419.Graham, D. E., R. Overbeek, G. J. Olsen, and C. R. Woese. 2000. An archaeal genomic signature. Proceedings of The National Academy of Sciences (U.S.A.) 97:3304-3308.Gray, M. W., G. Burger, and B. F. Lang. 1999. Mitochondrial evolution. Science 283:1476-1481.Gray, M.W. and Doolittle, W.F. (1982) Has the endosymbiont hypothesis been proven? Microbiol Rev., 46, 1-42.Gray, M.W., Burger, G. and Lang, B.F. (1999) Mitochondrial evolution. Science, 283, 1476-1481.Gray, M.W., Lang, B.F. and Burger, G. (2004) Mitochondria of protists. Annu. Rev. Genet., 38, 477-524.Gribaldo, S. and P. Cammarano. 1998. The root of the universal tree of life inferred from anciently duplicated genes encoding components of the protein-targeting machinery. Journal of Molecular Evolution 47:508-516.Griffiths, E. and R. S. Gupta. 2004. Signature sequences in diverse proteins provide evidence for the late divergence of the Order Aquificales. Int Microbiol. 7:41-52.Grosberg, R.K. and Strathmann, R.R. (2007) The evolution of multicellularity: a minor major transition? Annual Review of Ecology, Evolution, and Systematics, 38, 621-654.Gruber, T. M. and D. A. Bryant. 1997. Molecular systematic studies of eubacteria, using sigma(70)-type sigma factors of group 1 and group 2. Journal of Bacteriology 179:1734-1747.Gupta, R. S. 1997. Protein phylogenies and signature sequences: Evolutionary relationships within prokaryotes and between prokaryotes and eukaryotes. Antonie van Leeuwenhoek International Journal of General and Molecular Microbiology 72:49-61.Gupta, R. S. 1998. Protein phylogenies and signature sequences: A reappraisal of evolutionary relationships among archaebacteria, eubacteria, and eukaryotes. Microbiology and Molecular Biology Reviews 62:1435-1491.Gupta, R. S. 1998. What are archaebacteria: Life's third domain or monoderm prokaryotes related to Gram-positive bacteria? A new proposal for the classification of prokaryotic organisms. Molecular Microbiology 29:695-707.Gupta, R. S. 2000. The phylogeny of proteobacteria: relationships to other eubacterial phyla and eukaryotes. FEMS Microbiology Reviews 24(4):367-402.Gupta, R. S. 2004. The phylogeny and signature sequences characteristics of Fibrobacteres, Chlorobi, and Bacteroidetes. Critical Reviews in Microbiology 30(2):123-143.Gupta, R. S. and G. B. Golding. 1993. Evolution of HSP70 gene and its implications regarding relationships between archaebacteria, eubacteria, and eukaryotes. Journal of Molecular Evolution 37:573-582.Gupta, R. S., T. Mukhtar, and B. Singh. 1999. Evolutionary relationships among photosynthetic prokaryotes (Heliobacterium chlorum, Chloroflexus aurantiacus, cyanobacteria, Chlorobium tepidum and proteobacteria): implications regarding the origin of photosynthesis. Molecular Microbiology 32:893-906.Gupta, R.S. and E. Griffiths. 2002. Critical issues in bacterial phylogeny. Theoretical Population Biology 61(4):423-434.Gupta, R.S., K. Bustard, M. Falah, D. Singh. 1997. Sequencing of heat shock protein 70 (DnaK) homologs from Deinococcus proteolyticus and Thermomicrobium roseum and their integration in a protein-based phylogeny of prokaryotes. Journal of Bacteriology 179:345-357.Hackett, J.D., Yoon, H.S., Li, S., Reyes-Prieto, A., Rummele, S.E. and Bhattacharya, D. (2007) Phylogenomic analysis supports the monophyly of cryptophytes and haptophytes and the association of rhizaria with chromalveolates. Mol. Biol. Evol., 24, 1702-1713.Haen, K. M., B. F. Lang, S. A. Pomponi, and D. V. Lavrov. 2007. Glass sponges and bilaterian animals share derived mitochondrial genomic features: a common ancestry or parallel evolution? Molecular Biology and Evolution 24(7):1518 - 1527.Hagopian, J.C., Reis, M., Kitajima, J.P., Bhattacharya, D. and de Oliveira, M.C. (2004) Comparative analysis of the complete plastid genome sequence of the red alga Gracilaria tenuistipitata var. liui provides insights into the evolution of rhodoplasts and their relationship to other plastids. J. Mol. Evol., 59, 464-477.Halanych, K. 2004. The new view of animal phylogeny. Annual Review of Ecology, Evolution, and Systematics 35:229-256.Hampl, V., Horner, D.S., Dyal, P., Kulda, J., Flegr, J., Foster, P.G., and Embley, T.M. (2005) Inference of the phylogenetic position of oxymonads based on nine genes: Support for Metamonada and Excavata. Molecular Biology and Evolution, 22(12), 2508-2518.Hanelt B., D. VanSchyndel, C. M. Adema, L. A. Lewis, E. S. Loker. 1996. The phylogenetic position of Rhopalura ophiocomae (Orthonectida) based on 18S ribosomal DNA sequence analysis. Molecular Biology and Evolution 13:1187-1191.Hanson, E.D. (1977) The Origin and Early Evolution of Animals. Wesleyan University Press, Middletown, Conn.Harper, J.T. and Keeling, P.J. (2003) Nucleus-encoded, plastid-targeted glyceraldehyde-3-phosphate dehydrogenase (GAPDH) indicates a single origin for chromalveolate plastids. Mol. Biol. Evol., 20, 1730-1735.Hashimoto, T., Nakamura, Y., Kamaishi, T., and Hasegawa, M. (1997) Early evolution of eukaryotes inferred from protein phylogenies of translation elongation factors 1 alpha and 2. Archiv für Protistenkunde, 148, 287-295.Hendrix, R. W. 1999. The long evolutionary signNow of viruses. Current Biology 9:R914-R917.Hendrix, R. W., J. G. Lawrence, G. F. Hatfull, and S. Casjens. 2000. The origins and ongoing evolution of viruses. Trends in Microbiology 8:504-508.Hendrix, R. W., M. C. M. Smith, R. N. Burns, M. E. Ford, and G. F. Hatfull. 1999. Evolutionary relationships among diverse bacteriophages and prophages: all the world's a phage. Proceedings of the National Academy of Sciences (USA) 96:2192-2197.Herrmann, K.M. and Weaver, L.M. (1999) The Shikimate Pathway. Annu. Rev. Plant Physiol. Plant Mol. Biol., 50, 473-503.Hilario, E. and J. P. Gogarten. 1993. Horizontal transfer of ATPase genes: The tree of life becomes a net of life. Biosystems 31:111-119.Hirt, R.P. and Horner, D. (eds.) (2004) Organelles, Genomes and Eukaryote Evolution. Taylor & Francis, London.Hirt, R.P., Healy, B., Vossbrinck, C.R.,Canning, E.U., and Embley, T. M. (1997) A mitochondrial Hsp70 orthologue in Vairimorpha necatrix: Molecular evidence that microsporidia once contained mitochondria. Current Biology, 7, 995-998.Hirt, R.P., Logsdon, Jr., J.M., Healy, B., Dorey, M.W., Doolittle, W.F., and Embley, T.M. (1999) Microsporidia are related to fungi: evidence from the largest subunit of RNA polymerase II and other proteins. Proceedings of the National Academy of Sciences (USA), 96, 580-585.Hoppenrath, M. and Leander, B.S. (2006) Dinoflagellate, euglenid or cercomonad? The ultrastructure and molecular phylogenetic position of Protaspis grandis n. sp. J. Eukaryot. Microbiol., 53, 327-342.Huang, J., Xu, Y. and Gogarten, J.P. (2005) The presence of a haloarchaeal type tyrosyl-tRNA synthetase marks the opisthokonts as monophyletic. Mol. Biol. Evol., 22, 2142-2146.Huang, W. M. 1996. Bacterial diversity based on type II DNA topoisomerase genes. Annual Review of Genetics 30:79-107.Huang, Y. P. and J. Ito. 1999. DNA polymerase C of the thermophilic bacterium Thermus aquaticus: Classification and phylogenetic analysis of the family C DNA polymerases. Journal of Molecular Evolution 48:756-769.Huber, H., M. J. Hohn, R. Rachel, T. Fuchs, V. C. Wimmer, and K. O. Stetter. 2002. A new phylum of Archaea represented by a nanosized hyperthermophilic symbiont. Nature 417:63-67.Hugenholtz, P., B. M. Goebel, and N. R. Pace. 1998. Impact of culture-independent studies on the emerging phylogenetic view of bacterial diversity. Journal of Bacteriology 180:4765-4774.Iwabe, N., K.-I. Kuma, M. Hagesawa, S. Osawa, T. Miyata. 1989. Evolutionary relationship of archaebacteria, eubacteria, and eukaryotes inferred from phylogenetic trees of duplicated genes. Proceedings of the Natural Academy of Sciences (USA) 86:9355-9359.Iyer, L. M., L. Aravind, and E. V. Koonin. 2001. Common Origin of Four Diverse Families of Large Eukaryotic DNA Viruses. Journal of Virology 75(23):11720-11734.Jeffares, D. C., A. M. Poole, and D. Penny. 1998. Relics from the RNA world. Journal of Molecular Evolution 46:18-36.Jenner, R. A. 2004. The scientific status of metazoan cladistics: why current research practice must change. Zoologica Scripta 33(4):293-310.Jenner, R. A. 2004. When molecules and morphology clash: Reconciling conflicting phylogenies of the Metazoa by considering secondary character loss. Evolution & Development 6(5):372-378.Jenner, R. A. and D. T. J. Littlewood. 2008. Problematica old and new. Philosophical Transactions of the Royal Society Series B 363(1496):1503-1512.Jenner, R.A. and Schram, F.R. (1999) The grand game of metazoan phylogeny: rules and strategies. Biological Reviews, 74, 121-142.Jiménez-Guri, E., H. Philippe, B. Okamura, P. W. H. Holland. 2007. Buddenbrockia is a cnidarian worm. Science 317(5834):116-118.Johnson, M.D., Oldach, D., Delwiche, C.F. and Stoecker, D.K. (2007) Retention of transcriptionally active cryptophyte nuclei by the ciliate Myrionecta rubra. Nature, 445, 426-428.Kamaishi, T., Hashimoto, T., Nakamura, Y., Nakamura, F., Murata, S., Okada, N., Okamoto, K., and Hasegawa, M. (1996) Protein phylogeny of translation elongation factor EF-1alpha suggests microsporidians are extremely ancient eukaryotes. Journal of Molecular Evolution, 42, 257-263.Kamm, K., B. Schierwater, W. Jakob, S. L. Dellaporta, and D. J. Miller. 2006. Axial patterning and diversification in the Cnidaria predate the Hox system. Current Biology 16:920-926.Kandler, O. 1994. Cell wall biochemistry and three-domain concept of life. Systematic and Applied Microbiology 16:501-509.Katz, L. A. 1998. Changing perspectives on the origin of eukaryotes. Trends in Ecology and Evolution 13:493-497.Katz, L. A. 1999. The tangled web: gene genealogies and the origin of eukaryotes. Am. Nat. 154 (suppl.):S137-S145.Katz, L.A. (1998) Changing perspectives on the origin of eukaryotes. Trends Ecol. Evol., 13, 493-497.Katz, L.A. (1999) The tangled web: gene genealogies and the origin of eukaryotes. American Naturalist, 154(suppl.), S137-S145.Keeling, P.J. (1998) A kingdom's progress: Archezoa and the origin of eukaryotes. BioEssays, 20, 87-95.Keeling, P.J. (2001) Foraminifera and Cercozoa are related in actin phylogeny: two orphans find a home? Mol. Biol. Evol., 18, 1551-1557.Keeling, P.J. (2004) The diversity and evolutionary history of plastids and their hosts. Am. J. Bot., 91, 1481-1493.Keeling, P.J. (2009) Chromalveolates and the evolution of plastids by secondary endosymbiosis. J. Eukaryot Microbiol., in press.Keeling, P.J. and Doolittle, W.F. (1996) Alpha-tubulin from early-diverging eukaryotic lineages and the evolution of the tubulin family. Molecular Biology and Evolution, 13, 1297-1305.Keeling, P.J. and McFadden, G.I. (1998) Origins of microsporidia. Trends Microbiol., 6, 19-23.Keeling, P.J. and Palmer, J.D. (2000) Phylogeny - Parabasalian flagellates are ancient eukaryotes. Nature, 405, 635-637.Keeling, P.J., Burger, G., Durnford, D.G., Lang, B.F., Lee, R.W., Pearlman, R.E., Roger, A.J. and Gray, M.W. (2005) The tree of eukaryotes. Trends Ecol. Evol., 20, 670-676.Keeling, P.J., Luker, M.A., and Palmer, J.D. (2000) Evidence from beta-tubulin phylogeny that microsporidia evolved from within the fungi. Molecular Biology and Evolution, 17, 23-31.Kim, J., W. Kim, and C. W. Cunningham. 1999. A new perspective on lower metazoan relationships from 18S rDNA sequences. Molecular Biology and Evolution 16:423-427.Kim, J.,Kim, W., and Cunningham, C.W. (1999) A new perspective on lower metazoan relationships from 18S rDNA sequences. Molecular Biology and Evolution, 16, 423-427.Kjems, J., N. Larsen, J. Z. Dalgaard, R. A. Garrett and K. O. Stetter. 1992. Phylogenetic relationships amongst the hyperthermophilic Archaea determined from partial 23S rRNA gene sequences. Systematic and Applied Microbiology 15(2): 203-208.Klenk, H. P., C. Schleper, V. Schwass and R. Brudler. 1993. Nucleotide sequence, transcription and phylogeny of the gene encoding the superoxide dismutase of Sulfolobus acidocaldarius. Biochimica Et Biophysica Acta 1174(1): 95-98.Knoll, A. H. and S. B. Carroll. 1999. Early animal evolution: emerging views from comparative biology and geology. Science 284:2129-2137.Knoll, A.H. (1992) The early evolution of eukaryotes: a geological perspective. Science, 256, 622-627.Koonin, E. V., A. R. Mushegian, M. Y. Galperin, and D. R. Walker. 1997. Comparison of archaeal and bacterial genomes: computer analysis of protein sequences predicts novel functions and suggests a chimeric origin for the archaea. Molecular Microbiology 25:619-637.Kruse, M., S. P. Leys, I. M. Mueller, and W. E. G. Mueller. 1998. Phylogenetic position of the hexactinellida within the phylum porifera based on the amino acid sequence of the protein kinase C from Rhabdocalyptus dawsoni. Journal of Molecular Evolution 46:721-728.Kumar, S. and Rzhetsky, A. (1996) Evolutionary relationships of eukaryotic kingdoms. Journal of Molecular Evolution, 42, 183-193.Kunisawa, T. 2006. Dichotomy of major bacterial phyla inferred from gene arrangement comparisons. J. of Theor. Biol. 239:367-375.Kyrpides, N. C. and C. A. Ouzounis. 1999. Transcription in Archaea. Proceedings of The National Academy of Sciences (U.S.A.) 96:8545-8550.Kyrpides, N. C. and G. J. Olsen. 1999. Archaeal and bacterial hyperthermophiles: horizontal gene exchange or common ancestry? Trends in Genetics 15:298-299.Lake, J. A. 1990. Archaebacterial or eocyte tree? Nature 343:418-419.Lake, J. A. and M. C. Rivera. 1996. The prokaryotic ancestry of eukaryotes. Pages 87-108 in Evolution of Microbial Life (D. McL. Roberts, P. Sharp, G. Alderson, and M. Collins, eds.) Symposium 54. Society for General Microbiology. Cambridge University Press, Cambridge.Lake, J. A., M. W. Clark, E. Hendeson, S. P. Fay, M. Oakes, A. Scheinman, J. P. Thornber and R. A. Mah. 1985. Eubacteria, halobacteria and the origin of photosynthesis: The photocytes. Proceedings of the National Academy of Sciences (USA) 82:3716-3720.Lake, J.A. and Rivera, M.C. (1994) Was the nucleus the first endosymbiont? Proceedings of the National Academy of Sciences (USA), 91, 2880-2881.Lake, J.A., E. Henderson, M. Oakes, M.W. Clark. 1984. Eocytes: a new ribosome structure indicates a kingdom with close relationship to eukaryotes. Proceedings of the National Academy of Sciences (USA) 81:3786-3790.Lang, B.F., Burger, G., O'Kelly, C. J., Cedergren, R., Golding, G. B., Lemieux, C., Sankoff, D., Turmel, M., and Gray, M. W. (1997) An ancestral mitochondrial DNA resembling a eubacterial genome in miniature. Nature, 387, 493-497.Lang, B.F., Gray, M.W. and Burger, G. (1999) Mitochondrial genome evolution and the origin of eukaryotes. Annu. Rev. Genet., 33, 351-397.Lawrence, J. G. and H. Ochman. 1998. Molecular archaeology of the Escherichia coli genome. Proceedings of the National Academy of Sciences of the United States of America 95:9413-9417.Lawrence, J. G., G. F. Hatfull, and R. W. Hendrix. 2002. Imbroglios of viral taxonomy: Genetic exchange and failings of phenetic approaches. Journal of Bacteriology 184(17):4891-4905.Lawson, F. S., R. L. Charlebois, and J.-A. R. Dillon. 1996. Phylogenetic analysis of carbamoylphosphate synthetase genes: complex evolutionary history includes an internal duplication within a gene which can root the Tree of Life. Molecular Biology and Evolution 13:970-977.Leander, B.S. (2004) Did trypanosomatid parasites have photosynthetic ancestors? Trends Microbiol., 12, 251-258.Leander, B.S. (2008) A hierarchical view of convergent evolution in microbial eukaryotes. J. Eukaryot. Microbiol., 55, 59-68.Leander, B.S. (2008) Different modes of convergent evolution reflect phylogenetic distances. Trends Ecol. Evol., 23, 481-482.Leander, B.S. and Keeling, P.J. (2003) Morphostasis in alveolate evolution. Trends Ecol. Evol., 18, 395-402.Leander, B.S. and Keeling, P.J. (2004) Early evolutionary history of dinoflagellates and apicomplexans (Alveolata) as inferred from hsp90 and actin phylogenies. J. Phycol., 40, 341-350.Leipe, D., Gunderson, J.H., Nerad, T.A., and Sogin, M. L. (1993) Small subunit ribosomal RNA of Hexamita inflata and the quest for the first branch in the eukaryotic tree. Mol. Biochem. Parasitol., 59, 41-48.Levin, H. L. 1999. Ancient Invertebrates and Their Living Relatives. Prentics Hall, Upper Saddle River, New Jersey.Liao, D. and P. P. Dennis. 1994. Molecular phylogenies based on ribosomal protein L11, L1, L10, and L12 sequences. Journal of Molecular Evolution 38:405-419.Lipscomb D.L., Farris, J.S., Kallersjo, M., and Tehler, A. (1998) Support, ribosomal sequences and the phylogeny of the eukaryotes. Cladistics, 14, 303-338.Liu, R. and H. Ochman. 2007. Stepwise formation of the bacterial flagellar system. Proceedings of the National Academy of Sciences of the United States of America 104(17):7116-7121.Longet, D., Archibald, J.M., Keeling, P.J. and Pawlowski, J. (2003) Foraminifera and Cercozoa share a common origin according to RNA polymerase II phylogenies. Int. J. Syst. Evol. Microbiol., 53, 1735 - 1739.Lopez, P., P. Forterre, and H. Philippe. 1999. The root of the tree of life in the light of the covarian model. Journal of Molecular Evolution 49:496-508.Lovisolo, O., R. Hull, and O. Rösler. 2003. Coevolution of viruses with hosts and vectors and possible paleontology. Advances in Virus Research 62:325-379.Ludwig, W., J. Neumaier, N. Klugbauer, E. Brockmann, C. Roller, S. Jilg, K. Reetz, I. Schachtner, A. Ludvigsen, M. Bachleitner, U. Fischer, and K. H. Schleifer. 1993. Phylogenetic relationships of Bacteria based on comparative sequence analysis of elongation factor Tu and ATP-synthase beta-subunit genes. Antonie van Leeuwenhoek International Journal of General and Molecular Microbiology 64:285-305.Ludwig, W., O. Strunk, S. Klugbauer, N. Klugbauer, M. Weizenegger , J. Neumaier, M. Bachleitner, and K. H. Schleifer. 1998. Bacterial phylogeny based on comparative sequence analysis. Electrophoresis 19:554-568.Macario, A. J. L. and E. C. de Macario. 1999. The archaeal molecular chaperone machine: Peculiarities and paradoxes. Genetics 152:1277-1283.Makarova, K. S., L. Aravind, M. Y. Galperin, N. V. Grishin, R. L. Tatusov, Y. I. Wolf, and E. V. Koonin. 1999. Comparative genomics of the archaea (Euryarchaeota): Evolution of conserved protein families, the stable core, and the variable shell. Genome Research 9:608-628.Maldonado, M. (2004) Choanoflagellates, choanocytes, and animal multicellularity. Invertebrate Biology, 123, 1-22.Margulis, L. (1970) Origin of Eukaryotic Cells. Yale University Press.Margulis, L. (1981) Symbiosis in cell evolution. W. H. Freeman and Co., San Francisco.Margulis, L. 1996. Archaeal-eubacterial mergers in the origin of Eukarya: phylogenetic classification of life. Proceedings of the Natural Academy of Sciences (USA) 92:1071-1076.Margulis, L., Chapman, M., Guerrero, R., and Hall, J. (2006) The last eukaryotic common ancestor (LECA): Acquisition of cytoskeletal motility from aerotolerant spirochetes in the Proterozoic Eon. Proceedings of the National Academy of Sciences (USA), 103(35), 13080-13085.Margulis, L., Corliss, J.O., Melkonian, M., and Chapman, D.J. 1990. Handbook of Protoctista. Jones and Bartlett Publishers, Boston.Martin W. and M. Müller. 1998. The hydrogen hypothesis for the first eukaryote. Nature 392:37-41.Martin, W. 1999. Mosaic bacterial chromosomes: a challenge on route to a tree of genomes. BioEssays 21:99-104.Martindale, M. Q., J. R. Finnerty, and J. Q. Henry. 2002. The Radiata and the evolutionary origins of the bilaterian body plan. Molecular Phylogenetics and Evolution 24:358-365.Matte-Tailliez, O., C. Brochier, P. Forterre, and H. Philippe. 2002. Archaeal phylogeny based on ribosomal proteins. Molecular Biology and Evolution 19:631-639.Matus, D. Q., K. Pang, H. Marlow, C. W. Dunn, G. H. Thomsen, and M. Q. Martindale. 2006. Molecular evidence for deep evolutionary roots of bilaterality in animal development. Proceedings of the National Academy of Sciences (USA) 103(30):11195-11200.McClendon, J. H. 1999. The origin of life. Earth-Science Reviews 47:71-93.McCormack, G. P. and J. P. Clewley. 2002. The application of molecular phylogenetics to the analysis of viral genome diversity and evolution. Reviews in Medical Virology 12(4):221-238.McFadden, G.I. (1999) Endosymbiosis and evolution of the plant cell. Curr. Opin. Plant. Biol., 2, 513-519.McFadden, G.I., Gilson, P.R., Douglas, S.E., Cavalier-Smith, T., Hofmann, C.J. and Maier, U.G. (1997) Bonsai genomics: sequencing the smallest eukaryotic genomes. Trends Genet., 13, 46-49.Medina, M., A. G. Collins, J. D. Silberman, and M. L. Sogin. 2001. Evaluating hypotheses of basal animal phylogeny using complete sequences of large and small subunit rRNA. Proceedings of the National Academy of Sciences (U.S.A.) 98:9707-9712.Mendoza, L., Taylor, J.W., and Ajello, L. (2002) The class mesomycetozoea: a heterogeneous group of microorganisms at the animal-fungal boundary. Annual Review of Microbiology, 56, 315-44.Mindell, D. P., J. S. Rest, and L. P. Villarreal. 2004. Viruses and the tree of life. Pp. 107-118 in Cracraft, J. and M. J. Donoghue (eds.), Assembling the Tree of Life. Oxford University Press, New York.Minelli, A. 2007. Invertebrate taxonomy and evolutionary developmental biology. Pages 55-60 in: Zhang, Z.-Q. & Shear, W.A., eds. Linnaeus Tercentenary: Progress in Invertebrate Taxonomy. Zootaxa 1668:1-766.Minge, M.A., Silberman, J.D., Orr, R.J., Cavalier-Smith, T., Shalchian-Tabrizi, K., Burki, F., Skjaeveland, A. and Jakobsen, K.S. (2008) Evolutionary position of breviate amoebae and the primary eukaryote divergence. Proc. Biol. Sci., 276, 597-604.Monteiro, A. S., B. Okamura, and P. W. H. Holland. 2002. Orphan worm finds a home: Buddenbrockia is a Myxozoan. Molecular Biology and Evolution 19:968-971.Moran, N. and P. Baumann. 1994. Phylogenetics of cytoplasmically inherited microorganisms of arthropods. Trends in Ecology and Evolution 9:15-20.Moreira, D. and P. Lopez-Garcia. 1998. Symbiosis between methanogenic archaea and delta-proteobacteria as the origin of eukaryotes: the syntrophic hypothesis. Journal of Molecular Evolution 47:517-530.Moreira, D., Le Guyader, H. and Phillippe, H. (2000) The origin of red algae and the evolution of chloroplasts. Nature, 405, 69-72.Moreira, D., von der Heyden, S., Bass, D., Lopez-Garcia, P., Chao, E. and Cavalier-Smith, T. (2007) Global eukaryote phylogeny: Combined small- and large-subunit ribosomal DNA trees support monophyly of Rhizaria, Retaria and Excavata. Mol. Phylogenet. Evol., 44, 255-266.Morin, L. (2000) Long branch attraction effects and the status of "basal eukaryotes": Phylogeny and structural analysis of the ribosomal RNA gene cluster of the free-living diplomonad Trepomonas agilis. Journal of Eukaryotic Microbiology, 47, 167-177.Morris, P.J. (1993) The developmental role of the extracellular matrix suggests a monophyletic origin of the Kingdom Animalia. Evolution, 47, 152-165.Müller, M. (1993) The hydrogenosome. J. Gen. Microbiol., 139, 2879-2889.Narbonne, G.M. (2004) Modular construction of early Ediacaran complex life forms. Science, 305(5687), 1141-1144.Nealson, K. H. and P. G. Conrad. 1999. Life: past, present and future. Philosophical Transactions of the Royal Society of London Series B 354:1923-1939.Nielsen, C. 2001. Animal Evolution: Interrelationships of the Living Phyla. Second Edition. Oxford University Press, Oxford.Nielsen, C., N. Scharff, and D. Eibye-Jacobsen. 1996. Cladistic analyses of the animal kingdom. Biological Journal of the Linnean Society 57:385-410.Nikolaev, S.I., Berney, C., Fahrni, J.F., Bolivar, I., Polet, S., Mylnikov, A.P., Aleshin, V.V., Petrov, N.B. and Pawlowski, J. (2004) The twilight of Heliozoa and rise of Rhizaria, an emerging supergroup of amoeboid eukaryotes. Proc. Natl. Acad. Sci. USA, 101, 8066-8071.Nowack, E.C., Melkonian, M. and Glockner, G. (2008) Chromatophore genome sequence of Paulinella sheds light on acquisition of photosynthesis by eukaryotes. Curr. Biol., 18, 410-418.Nozaki, H., Iseki, M., Hasegawa, M., Misawa, K., Nakada, T., Sasaki, N., and Watanabe, M. (2007) Phylogeny of primary photosynthetic eukaryotes as deduced from slowly evolving nuclear genes. Molecular Biology and Evolution, 24, 1592-1595.Nozaki, H., Matsuzaki, M., Takahara, M., Misumi, O., Kuroiwa, H., Hasegawa, M., Shin-i, T., Kohara, Y., Ogasawara, N., and Kuroiwa, T. (2003) The phylogenetic position of red algae revealed by multiple nuclear genes from mitochondria-containing eukaryotes and an alternative hypothesis on the origin of plastids. Journal of Molecular Evolution, 56(4), 485-497.Ochman, H., S. Elwyn, and N. A. Moran. 1999. Calibrating bacterial evolution. Proceedings of the National Academy of Sciences of the United States of America 96:12638-12643.Okamoto, N. and Inouye, I. (2005) A secondary symbiosis in progress? Science, 310, 287.Okamura, B., A. Curry, T. S. Wood, and E. U. Canning. 2002. Ultrastructure of Buddenbrockia identifies it as a myxozoan and verifies the bilaterian origin of the Myxozoa. Parasitology 124:215-223.Olsen, G. J. and C. R. Woese. 1993. Ribosomal RNA: a key to phylogeny. FASEB Journal 7:113-23.Olsen, G. J., C. R. Woese, and R. Overbeek. 1994. The winds of (evolutionary) change: breathing new life into microbiology. Journal of Bacteriology 176:1-6.Pace, N. R. 1997. A molecular view of microbial diversity and the biosphere. Science 276:734-740.Pace, N. R. 1999. Microbial ecology and diversity. ASM News 65:328-333.Patron, N.J., Inagaki, Y. and Keeling, P.J. (2007) Multiple gene phylogenies support the monophyly of cryptomonad and haptophyte host lineages. Curr. Biol., 17, 887-891.Patron, N.J., Rogers, M.B. and Keeling, P.J. (2004) Gene replacement of fructose-1,6-bisphosphate aldolase (FBA) supports a single photosynthetic ancestor of chromalveolates. Eukaryot. Cell, 3, 1169-1175.Patterson, D.J. (1994) Protozoa: Evolution and Systematics. Pages 1-14 in Progress in Protozoology. Proceedings of the IX International Congress of Protozoology, Berlin (1993) (K. Hausmann and N. Hülsmann, eds.) Gustav Fischer Verlag, Stuttgart, Jena, New York.Patterson, D.J. (1999) The diversity of eukaryotes. American Naturalist, 154(suppl.), S96-S124.Patterson, D.J. and Sogin, M.L. (1992) Eukaryote origins and protistan diversity. Pages 13-46 in The Origin and Evolution of Prokaryotic and Eukaryotic Cells (H. Hartman and K. Matsuno, eds.) World Scientific Pub. Co. NJ.Pennisi, E. 1998. Genome data shake the tree of life. Science 280:672-674.Pennisi, E. 1999. Is it time to uproot the tree of life? Science 284:1305-1307.Penny, D. and A. Poole. 1999. The nature of the last universal common ancestor. Current Opinion in Genetics and Development 9:672-677.Peterson, K. J. and D. J. Eernisse. 2001. Animal phylogeny and the ancestry of bilaterians: inferences from morphology and 18S rDNA gene sequences. Evolution & Development 3:170-205.Peterson, K. J. and E. H. Davidson. 2000. Regulatory evolution and the origin of bilaterians. Proceedings of the National Academy of Sciences (U.S.A.) 97:4430-4433.Peterson, K. J. and N. J. Butterfield. 2005. Origin of the Eumetazoa: Testing ecological predictions of molecular clocks against the Proterozoic fossil record. Proceedings of the National Academy of Sciences (USA) 102(27):9547-9552.Peterson, K. J., J. A. Cotton, J. G. Gehling and D. Pisani. 2008. The Ediacaran emergence of bilaterians: congruence between the genetic and the geological fossil records. Philosophical Transactions of the Royal Society Series B 363(1496):1435-1443.Peterson, K. J., J. B. Lyons, K. S. Nowak, C. M. Takacs, M. J. Wargo and M. A. McPeek. 2004. Estimating metazoan divergence times with a molecular clock. Proceedings of the National Academy of Sciences (USA) 101(17):6536-6541.Philip, G.K., Creevey, C.J., and McInerney, J.O. (2005) The Opisthokonta and the Ecdysozoa may not be clades: Stronger support for the grouping of plant and animal than for animal and fungi and stronger support for the Coelomata than Ecdysozoa. Molecular Biology and Evolution, 22(5), 1175.1184.Philippe, H. and Adoutte, A. (1998) The molecular phylogeny of Eukaryota: solid facts and uncertainties. Pages 25-56 in Evolutionary Relationships among Protozoa (G. H. Coombs, K. Vickerman, M. A. Sleigh, and A. Warren, eds.) Chapman & Hall, London.Philippe, H. and Germot, A. (2000) Phylogeny of eukaryotes based on ribosomal RNA: Long-branch attraction and models of sequence evolution. Molecular Biology and Evolution, 17, 830-834.Philippe, H. and M. J. Telford. 2006. Large-scale sequencing and the new animal phylogeny. Trends in Ecology & Evolution 21(11):614-620.Philippe, H. and P. Forterre. 1999. The rooting of the universal tree of life is not reliable. Journal of Molecular Evolution 49:509-523.Philippe, H., Lopez, P., Brinkmann, H., Budin, K., Germot, A., Laurent, J., Moreira, D., Muller, M., and Le Guyader, H. (2000) Early-branching or fast-evolving eukaryotes? An answer based on slowly evolving positions. Proceedings of the Royal Society of London Series B, 267, 1213-1221.Philippe, H., Snell, E.A., Bapteste, E., Lopez, P., Holland, P.W.H., and Casane, D. (2004) Phylogenomics of eukaryotes: impact of missing data on large alignments. Molecular Biology and Evolution, 21(9), 1740-1752.Pierson, B. K. 1994. The emergence, diversification, and role of photosynthetic bacteria. Pages 161-180 in Early Life on Earth, Nobel Symposium No. 84 (Bengtson, S., ed.). Columbia University Press, New York.Polet, S., Berney, C., Fahrni, J. and Pawlowski, J. (2004) Small-subunit ribosomal RNA gene sequences of Phaeodarea challenge the monophyly of Haeckel's Radiolaria. Protist, 155, 53-63.Poole, A., D. Jeffares, and D. Penny. Early evolution: prokaryotes, the new kids on the block. BioEssays 21:880-889.Putnam, N. H., M. Srivastava, U. Hellsten, B. Dirks, J. Chapman, A. Salamov, A. Terry, H. Shapiro, E. Lindquist, V. V. Kapitonov, J. Jurka, G. Genikhovich, I. V. Grigoriev, S. M. Lucas, R. E. Steele, J. R. Finnerty, U. Technau, M. Q. Martindale, and D. S. Rokhsar. 2007. Sea anemone genome reveals ancestral eumetazoan gene repertoire and genomic organization. Science 317(5834):86-94.Ragan, M.A. and Gutell, R.R. (1995) Are red algae plants? Botanical Journal of the Linnean Society, 118, 81-105.Ragan, M.A., Goggin, C.L., Cawthorn, R.J., Cerenius, L., Jamieson, A.V., Plourde, S.M., Rand, T.G., Soderhall, K. and Gutell, R.R. (1996) A novel clade of protistan parasites near the animal-fungal divergence. Proc. Natl. Acad. Sci. USA, 93, 11907-11912.Rappé, M. S. and S. J. Giovannoni. 2003. The uncultured microbial majority. Annual Review of Microbiology 57:369-394.Rasmussen, B. 2000. Filamentous microfossils in a 3,235-million-year-old volcanogenic massive sulphide deposit. Nature 405:676-679.Reichert, A.S. and Neupert, W. (2004) Mitochondriomics or what makes us breathe. Trends Genet., 20, 555-562.Reyes-Prieto, A., Weber, A.P. and Bhattacharya, D. (2007) The origin and establishment of the plastid in algae and plants. Annu. Rev. Genet., 41, 147-168.Reysenbach1, A. L. and E. Shock. 2002. Merging genomes with geochemistry in hydrothermal ecosystems. Science 296:1077-1082.Ribeiro, S. and G. B. Golding. 1998. The mosaic nature of the eukaryotic nucleus. Molecular Biology and Evolution 15:779-788.Ribeiro, S. and Golding, G.B. (1998) The mosaic nature of the eukaryotic nucleus. Molecular Biology and Evolution, 15, 779-788.Rice, D.W. and Palmer, J.D. (2006) An exceptional horizontal gene transfer in plastids: gene replacement by a distant bacterial paralog and evidence that haptophyte and cryptophyte plastids are sisters. BMC Biol., 4, 31.Rice, G., L. Tang, K. Stedman, F. Roberto, J. Spuhler, E. Gillitzer, J. E. Johnson, T. Douglas, and M. Young. 2004. The structure of a thermophilic archaeal virus shows a double-stranded DNA viral capsid type that spans all domains of life. Proceedings of the National Academy of Sciences (USA) 101(20):7716-7720.Richards, T.A. and Cavalier-Smith, T. (2005) Myosin domain evolution and the primary divergence of eukaryotes. Nature, 436, 1113-1118.Richards, T.A. and van der Giezen, M. (2006) Evolution of the Isd11-IscS complex reveals a single alpha-proteobacterial endosymbiosis for all eukaryotes. Molecular Biology and Evolution, 23, 1341-1344.Rivera, M. C., R. Jain, J. E. Moore, and J. A. Lake. 1998. Genomic evidence for two functionally distinct gene classes. Proceedings of the National Academy of Sciences (USA) 95:6239-6244.Rivera, M. C., and J. A. Lake. 2004. The ring of life provides evidence for a genome fusion origin of eukaryotes. Nature 431:152-155.Robinson, R. 2005. Jump-starting a cellular world: Investigating the origin of life, from soup to networks. PLoS Biol 3(11): e396. http://biology.plosjournals.org/...Rodriguez-Ezpeleta, N., Brinkmann, H., Burey, S.C., Roure, B., Burger, G., Loffelhardt, W., Bohnert, H.J., Philippe, H. and Lang, B.F. (2005) Monophyly of primary photosynthetic eukaryotes: green plants, red algae, and glaucophytes. Curr Biol, 15, 1325-1330.Rodriguez-Ezpeleta, N., Brinkmann, H., Burger, G., Roger, A.J., Gray, M.W., Philippe, H. and Lang, B.F. (2007) Toward resolving the eukaryotic tree: the phylogenetic positions of jakobids and cercozoans. Curr. Biol., 17, 1420-1425.Roger, A.J. (1999) Reconstructing early events in eukaryotic evolution. Am. Nat., 154, S146-S163.Roger, A.J., Sandblom, O., Doolittle, W. F., and Philippe, H. (1999) An evaluation of elongation factor 1 alpha as a phylogenetic marker for eukaryotes. Molecular Biology and Evolution, 16, 218-233.Rogers, M.B., Gilson, P.R., Su, V., McFadden, G.I. and Keeling, P.J. (2007) The complete chloroplast genome of the chlorarachniophyte Bigelowiella natans: evidence for independent origins of chlorarachniophyte and euglenid secondary endosymbionts. Mol. Biol. Evol., 24, 54-62.Ruiz-Trillo, I., A. J. Roger, G. Burger, M. W. Gray, and B. F. Lang. 2008. A phylogenomic investigation into the origin of Metazoa. Molecular Biology and Evolution 25:664-672.Ruiz-Trillo, I., Lane, C.E., Archibald, J.M. and Roger, A.J. (2006) Insights into the evolutionary origin and genome architecture of the unicellular opisthokonts Capsaspora owczarzaki and Sphaeroforma arctica. J. Eukaryot. Microbiol., 53, 379-384.Ruppert, E. E., R. S. Fox, and R. D. Barnes. 2004. Invertebrate Zoology, a Functional Evolutionary Approach. 7th ed. Brooks/Cole-Thomson Learn, Belmont, CA.Sapp, J. 2005) Microbial Phylogeny and Evolution: Concepts and Controversies. Oxford University Press, New York.Schierwater, B., M. Eitel, W. Jakob, H.-J. Osigus, H. Hadrys, S. L. Dellaporta, S.-O. Kolokotronis, and R. DeSalle. 2009. Concatenated molecular and morphological analysis sheds light on early metazoan evolution and fuels a modern .Urmetazoon. hypothesis. PLoS Biol 7(1): e1000020. doi:10.1371/journal.pbio.1000020.Schlegel, M. (2003) Phylogeny of Eukaryotes recovered with molecular data: highlights and pitfalls. European Journal of Protistology, 39, 113-122.Schlegel, M., J. Lom, A. Stechmann, D. Bernhard, D. Leipe, I. Dykova, and M. L. Sogin. 1996. Phylogenetic analysis of complete small subunit ribosomal RNA coding region of Myxidium lieberkuehni: Evidence that Myxozoa are Metazoa and related to the Bilateria. Archiv für Protistenkunde 147:1-9.Schopf, J. W. 2006. Fossil evidence of Archaean life. Philos. T. R. Soc. B 361:869-85.Schütze J., Krasko, A., Custodio, M.R., Efremova, S.M., Müller, I.M. and Müller, W.E.G. (1999) Evolutionary relationships of Metazoa within the eukaryotes based on molecular data from Porifera. Proceedings of the Royal Society of London Series B, 266, 63-73.Sicheritz-Ponten, T., C. G. Kurland, and S. G. E. Andersson. 1998. A phylogenetic analysis of the cytochrome b and cytochrome c oxidase I genes supports an origin of mitochondria from within the Rickettsiaceae. Biochimica et Biophysica Acta Bioenergetics 1365:545-551.Siddall, M. E. and M. F. Whiting. 1999. Long-branch abstractions. Cladistics 15:9-24.Siddall, M. E., D. S. Martin, D. Bridge, S. S. Desser, and D. K. Cone. 1995. The demise of a phylum of protists: Phylogeny of myxozoa and other parasitic cnidaria. Journal of Parasitology 81:961-967.Siddall, M.E., Martin, D.S., Bridge, D., Desser, S.S., and Cone, D.K. (1995) The demise of a phylum of protists: phylogeny of Myxozoa and other parasitic Cnidaria. J. Parasitol., 81, 961-967.Simpson, A.G. (2003) Cytoskeletal organization, phylogenetic affinities and systematics in the contentious taxon Excavata (Eukaryota). Int. J. Syst. Evol. Microbiol., 53, 1759-1777.Simpson, A.G. and Patterson, D.J. (2001) On core jakobids and excavate taxa: the ultrastructure of Jakoba incarcerata. J. Eukaryot. Microbiol., 48, 480-492.Simpson, A.G. and Roger, A.J. (2002) Eukaryotic evolution: getting to the root of the problem. Curr Biol, 12, R691-693.Simpson, A.G., Inagaki, Y. and Roger, A.J. (2006) Comprehensive multigene phylogenies of excavate protists reveal the evolutionary positions of "primitive" eukaryotes. Mol. Biol. Evol., 23, 615-625.Simpson, A.G., Roger, A.J., Silberman, J.D., Leipe, D.D., Edgcomb, V.P., Jermiin, L.S., Patterson, D.J. and Sogin, M.L. (2002) Evolutionary history of "early-diverging" eukaryotes: the excavate taxon Carpediemonas is a close relative of Giardia. Mol. Biol. Evol., 19, 1782-1791.Simpson, A.G.B. and Patterson, D.J. (1999) The ultrastructure of Carpediemonas membranifera (Eukaryota) with reference to the "Excavate hypothesis". Eur. J. Protistol., 35, 353-370.Skophammer, R. G., C. W. Herbold, M. C. Rivera, J. A. Servin, and J. A. Lake. 2006. Evidence that the Root of the Tree of Life Is Not within the Archaea. Molecular Biology and Evolution 23(9):1648-1651.Smothers, J. F., C. D. von Dohlen, L. H. Smith, Jr., and R. D. Spall. 1994. Molecular evidence that the myxozoan protists are metazoans. Science 265:1719-1721.Smothers, J.F., van Dohlen, C.D., Smith, L.H., and Spall, R. D. (1994) Molecular evidence that the myxozoan protists are metazoans. Science, 265, 1719-1721.Sogin, M.L. (1989) Evolution of eukaryotic microorganisms and their small subunit ribosomal RNAs. Amer. Zool., 29, 487-499.Sogin, M.L. (1991) Early evolution and the origin of eukaryotes. Current Opinion in Genetics and Development, 1, 457-463.Sogin, M.L. and Silberman, J.D. (1998) Evolution of the protists and protistan parasites from the perspective of molecular systematics. International Journal of Parasitology, 28, 11-20.Sogin, M.L., Elwood, H.J. and Gunderson, J.H. (1986) Evolutionary diversity of eukaryotic small-subunit rRNA genes. Proc. Natl. Acad. Sci. USA, 83, 1383-1387.Sogin, M.L., Morrison, H.G., Hinkle, G., and Silberman, J. D. (1996) Ancestral relationships of the major eukaryotic lineages. Microbiologia SEM, 12, 17-28.Staley, J. T. and J. J. Gosink. 1999. Poles apart: Biodiversity and biogeography of sea ice bacteria. Annual Review of Microbiology 53:189-215.Stanier, R.Y. (1970) Some aspects of the biology of cells and their possible evolutionary significance. Symp. Soc. Gen. Mircrobiol., 20, 1-38.Stechmann, A. and Cavalier-Smith, T. (2002) Rooting the eukaryote tree by using a derived gene fusion. Science, 297, 89-91.Stechmann, A. and Cavalier-Smith, T. (2003) The root of the eukaryote tree pinpointed. Curr. Biol., 13, R665-666.Stechmann, A. and T. Cavalier-Smith. (2002) Rooting the eukaryote tree by using a derived gene fusion. Science 297:89-91.Steenkamp, E.T., Wright, J. and Baldauf, S.L. (2006) The protistan origins of animals and fungi. Mol. Biol. Evol., 23, 93-106.Stetter, K. O. 1996. Hypterthermophilic procaryotes. FEMS Microbiology Reviews 18:149-158.Stiller, J.W. and Hall, B.D. (1997) The origin of red algae: Implications for plastid evolution. Proceedings of the National Academy of Sciences (USA), 94, 4520-4525.Stiller, J.W. and Hall, B.D. (1999) Long-branch attraction and the rDNA model of early eukaryotic evolution. Molecular Biology and Evolution 16:1270-1279.Stiller, J.W., Duffield, E.C.S., and Hall, B.D. (1998) Amitochondriate amoebae and the evolution of DNA-dependent RNA polymerase II. Proceedings of the National Academy of Sciences (USA), 95, 11769-11774.Stiller, J.W., Riley, J
-
How can I apply for SBI PO online?
HOW TO APPLYCandidates can apply online only from 07.02.2017 to 06.03.2017 and no other mode of application will be accepted.(a) GUIDELINES FOR FILLING ONLINE APPLICATION:Candidates will be required to register themselves online through Bank's website www.sbi.co.in/careers or http://www.statebankofindia.com/.... After registration candidates are required to pay the requisite application fee through online mode by using debit card/ credit card/Internet Banking.Helpdesk: In case of any problem in filling up the form, payment of fee/intimation charges or receipt of Admission/call letter, queries may be made at telephone no. 022-22820427 (between 11:00 AM to 06:00 PM on working days) or lodge his/her query on IBPS Candidate Grievance Redressal System. Candidates are advised to not to forget to mention 'Recruitment of Probationary Officers in State Bank of India-2017' in the subject of the email.Pre-requisites for Applying Online: Candidates should have valid email ID which should be kept active till the declaration of results. It will help him/her in getting call letter/interview advices etc. By email.( b) PAYMENT OF FEES : [ONLINE MODE ONLY] :i. Candidates should first scan their photograph and signature as detailed under guidelines for scanning the photograph and signature.ii. Candidates to visit Bank's website http://www.statebankofindia.com/... or State Bank of India and open the appropriate Online Application Format, available in the current openings.iii. Fill the application carefully. Once the application is filled in completely, candidates should submit the data. In the event of candidates not being able to fill the data in one go, they can save the data already entered. When the data is saved, a provisional registration number and password will be generated by the system and displayed on the screen. Candidates should note down the registration number and password. They can reopen the saved data using registration number and password and edit the particulars, if needed. This facility will be available three times only. Once the application is filled in completely, candidates should submit the data. No change /edit will be allowed thereafter. The registration at this stage is provisional.iv. After ensuring the correctness of the particulars of the application form, candidates are required to pay fees through the payment gateway integrated with the application, following the instructions available on the screen.v. The payment can be made by using debit card/ credit card/ Internet Banking by providing information as asked on the screen. Transaction charges for online payment, if any, will be borne by the candidates.vi. On successful completion of the transaction, e-receipt and application form will be generated; whichmay be printed for record. The printout of the application form is not to be sent to the Bank.vii. If the online transaction is not successfully completed, please register again and make payment online.viii. There is also a provision to reprint the e-Receipt and Application form containing fee details, at later stage.Source: SBI
-
Why do people claim North Africa wasn't populated by negroids, but genetic says otherwise? Haplogroups E and L (including subcla
Because its not true. Further this is a pointless question and doesnt prove anything regarding skin tone or your physical appearance since the Y-chromosome does NOT carry one of the genes for skin color. The Y-chromosome is actually mostly a genetic desert. The two major genes for skin color are on Chromosome 5 and Chromosome 15. Y chromosomal haplogroups and mtDNA haplogroups are not directly related to the autosomal admixture (genetics)Either way haplogroup E is not “black” is this “black” here is a synonym for the negroid “race” and mtdna L is not indigenous to North Africa nor that it equals “Black” either otherwise this would imply that Negroids were involved in the ooa which is not supported by any evidence.“there is no evidence of any L-derived clade outside Africa with a similar coalescence age to that proposed [OOA] movement." Fregel et al 2013"M168 (CT) and L3 had African descendants" ...... "the Eurasian "Adam" M168 could have met his equivalent, the Eurasian Eve L3" Johnson; 2007Here's what anthropology and genetic experts claim :“Migration out of Africa happened perhaps 50,000, 70,000 or even 200,000 years ago, long before 2,000 BC, when the first proto-Bantu appeared. Bantu people settled throughout (subsaharan) Africa after 1,000 BC.. Early migrating Africans [OOA] … were certainly not Bantu.” – (Daems; 2007)‘’The notions that chimpanzees are the ancestors of humans and that Africans are the ancestral human populations are false: rather, modern Africans descend from a population that is ancestral to all humans."Tishkoff et al 2012Further the indigenous haplogroups, meaning derived from a non colonial population, meaning the first humans to step in North Africa were Y haplogroup E1b1b and mtdna U6 berber and M1a in the case of Egypt not L. Both lineages are indigenous to North Africa yet near eastern/Eurasian in origin.U6 is a sister of european u5 and basically a cro magnon lineage. Basal u6 has been found in Romania and migrated more than 30.000 years ago into North Africa.U6 is a non-African lineage and represents a back-migration to Africa. (Hodgson et al; 2014) Therefore, there is no reason to suppose a sub-Saharan origin of ancestral haplogroup U (Smith et al; 2013). "U6 traces the first detectable Paleolithic return to Africa of ancient Caucasoid lineages. It has been mostly found in Northwest Africa, with a global estimated age of 47,000 years reflecting an old human continuity in that rather isolated area." (Maca-Meyer et al ; 2001)"U6 is the sister of U5, the oldest haplogroup in Europe, up to 50,000 years. U5 and U6 have a common ancestor in the Near East from which these two branches diverged. - one populating the Northern Mediterranean coast, giving rise to U5 and the second colonizing the southern Mediterranean coast, giving rise to U6 in North Africa. U6 marks the signature of the return of modern man toward Africa around 39,000-52,000 years ago." (Rando et al 1998; Macaulay et al 1999; Maca-Meyer et al 2001; Maca-Meyer et al 2003) "The Iberomaurusian was initiated by an expansion of modern humans of ultimately Near Eastern, carrying mtDNA haplogroup U6, who had spread into Cyrenaïca ~35-45 ka and produced the Dabban industry. The link back to the Near East and the European Early Upper Palaeolithic (which likely has the same source) may explain the suggested skeletal similarities between the robust Iberomaurusian "Mechta-Afalou" burials and European Cro-Magnon remains, as well as the case for continuity of the bearers of the Iberomaurusian industry from Morocco with later northwest African populations suggested by the dental evidence Pereira et al 2010.The oldest E-M215 also known as E-M35 has been found in Morocco at taforalt.Carriers of mitochondrial DNA macrohaplogroup L3 basal lineages migrated back to Africa from Asia around 70,000 years agoCarriers of mitochondrial DNA macrohaplogroup L3 basal lineages migrated back to Africa from Asia around 70,000 years agoNorth Africa has not been populated by negroid Africans pre the trans saharan slave trade started by the arabs, 700 to 1400 years ago..“We estimate that a migration of western African origin into Morocco began about 40 generations ago (approximately 1,200 ya); a migration of individuals with Nilotic ancestry into Egypt occurred about 25 generations ago (approximately 750 ya). Our genomic data reveal an extraordinarily complex history of migrations, involving at least five ancestral populations, into North Africa”Source:Genomic Ancestry of North Africans Supports Back-to-Africa Migrationshttps://www.google.com/url?sa=t&...There is no support by those genetics that the indigenous people have been replaced..and they are not negroid, “Black” Africans nor arab nor european either. All previous genetic studies have demonstrated biological continuity in North Africa and hinted at the ancients having affinity with west Asians and Europeans and this affinity is NOT due to recent near eastern or european migrations."Indigenous North Africans are genetically quite distinct from sub-Saharan Africans (1), and this difference is reflected in their lighter skin and European/Middle Eastern physical features." Science 6 2007:Vol. 316. no. 5821, pp. 50 – 53"The Nile River delta population is mainly Caucasian in origin"(Herrera et al; 2004)The term Caucasian (which refers to the inhabitants of the Caucasus) has been previously used instead of Caucasoid. caucasoid does not equal EuropeanMummy portraits or Fayum mummy portraits is the modern term given to a type of naturalistic painted portraits on wooden boards attached to mummies from the Coptic period. They belong to the tradition of panel painting, one of the most highly regarded forms of art in the Classical world. In fact, the Fayum portraits are the only large body of art from that tradition to have survived.Mummy portraits have been found across Egypt, but are most common in the Fayum Basin, particularly from Hawara and Antinoopolis, hence the common name. "Faiyum Portraits" is generally thought of as a stylistic, rather than a geographic, description. While painted Cartonnage mummy cases date back to pharaonic times, the Faiyum mummy portraits were an innovation dating to the Coptic period on time of the Roman occupation of Egypt. They date to the Roman period, from the late 1st century BCE or the early 1st century CE onwards. It is not clear when their production ended, but recent research suggests the middle of the 3rd century. They are among the largest groups among the very few survivors of the highly prestigious panel painting tradition of the classical world, which was continued into Byzantine and Western traditions in the post-classical world, including the local tradition of Coptic iconography in Egypt.The portraits covered the faces of bodies that were mummified for burial.The dental morphology of the Roman-period Fayum mummies was compared with that of earlier Egyptian populations, and was found to be "much more closely akin" to that of ancient Egyptians than to Greeks or other European populationsThose people who make these “claims” are scientists. The people who do this research, base their lives around it. They aren't just coming up with this stuff, because they dislike people. If they did, their careers would be over, because it makes them unethical and unprofessional. All credibility gone. No more access to the sites. No more funding. In science,a theory is not guesswork,but something which takes facts we know to be verifiable, and uses evidence to explain those facts. This “claim” that north Africa has never been racially or genetically the same as sub saharan Africa is not just a theory made up by “people” nor that its a fallacy.. A theory is the final step that a hypothesis signNowes when the body of facts supports it.. The evidence is clear.Haplogroup E is Eurasian in origin according to Spencer Wells and Oppenheimer.Y-DNA E is descended from Non-African CT-168 population and all Haplogroups that are descended from CT-M168 are equally Eurasian** Phylo-genetically. CT-M168 could be seen as the Eurasian Adam - the great … great-grandfather of every non-African man alive today.*Y-DNA haplogroup E would appear to have arisen in Africa based on the concentration and variety of E subclades in that area today. But the fact that Haplogroup E is closely linked with Haplogroup D, which is not found in Africa, leaves open the possibility that E first arose in the Near or Middle East and was subsequently carried into Africa by a back migration.Carriers of mitochondrial DNA macrohaplogroup L3 basal lineages migrated back to Africa from Asia around 70,000 years ago“Non-African backflow in the Y locus includes E and could ultimately massively outnumber the original African Y throughout Africa, ranging from 59 per cent (rather higher than mtDNA figures cited above) in East Africa and 65–80% in South Africa, to well over 90 per cent in West Africa (data from Cruciani et al. [61])." (Oppenheimer; 2012)"All Y lineages that arose from the M168 lineage do not trace to an African origin. ...Some of the YAP insertion chromosomes without the M174 mutation signNowed the Mediterranean via Central Asia and gave rise to the E lineage with mutations at M40 and M96 (~31 000 years ago; Hammer et al. 1998). This E lineage back-migrated to Africa through the Levant as hypothesized by Hammer et al. (1997) and Altheide and Hammer (1997). It is also evident that haplotype E-M34 chromosomes were probably introduced into Ethiopia from the Near East (Cruciani et al. 2004). The hypothesis of a back migration from Asia to Africa is strongly supported by the current phylogeography of the Y-chromosome variation, because haplogroup K2 and paragroup R1b*, both belonging to the otherwise Asiatic macro haplogroup K, have only been observed at high frequencies in Africa (Cruciani et al. 2002; Luis et al. 2004). Thus the major sub-sets of Y lineages that arose from the M168 lineage do not trace to an African origin."Chandrasekar et al. 2007Regions near but external to northeast Africa, like the Levant or the southern Arabian Peninsula could have served as an incubator for the early diversification of non-African uniparental haplogroup varieties like Y chromosome DE-YAP, CF-P143* and mtDNA M and N molecular ancestors. These would have spread globally and diversified over time and space. This model would imply that both CF-P143 and the DE-YAP evolved nearby but outside Africa. One DE-YAP* ancestor would have spread to Asia and evolved to haplogroup D while another DE-YAP* returned to northeast Africa and evolved into hg E. (Abu-Amero; 2009)“The YAP insertion probably occurred on an Asian Y chromosome as long ago as 55 000 years. M168 chromosomes have given rise to the YAP insertion and M174 mutation in south Asia. Some of the YAP insertion chromosomes without the M174 mutation signNowed the Mediterranean via Central Asia and gave rise to the E lineage with mutations at M40 and M96 This E lineage back-migrated to Africa through the Levant.” (Chandrasekar et al; 2007)E1b1 is by far the lineage of greatest geographical distribution. It has two important sub-lineages, E1b1a and E1b1b. E1b1a is an African lineage that probably expanded from northern African to sub-Saharan and equatorial Africa with the Bantu agricultural expansion.“Black” or sub saharan E carriers are a mutation of Eurasian males are female pygmies. It was eurasians who migrated south. Their ancient eurasian admixture is from the remaining of the sahara original people known as the kiffians (a mechtoid sub-type) The Negroid gets its height from Eurasians. North Africans are in fact much older than “Black” Africans.Negroids radiated from a relatively small West African Pygmy population within the last 12,000 years (Coon, 1962, pp. 651-656; Spurdle et al., 1994; Watson et al., 1996).Black or negroid Africans are the youngest edition to the African continent that’s generally accepted to have formed somewhere in West-Central Africa through the mixture of archaic Africans (A, B and L0-L2 in terms of haplogroups) and proto-Eurasians (CT →E1a and R-V88 and the descendants of L3), and spread throughout Africa only in recent times during the bantu expansion/migration which occured within the last 5000 years when the sahara started to dry out. “black” Africans from Equatorial lands are also the only 'species' that carry the Muc7 "ghost /archaic gene"The paternal lineage of most sub saharan “black” Africans( incl “Black” -E carrying -Americans) is Eurasian is origin, which received minor neanderthal dna, and maternal pygmy which is mixed archaic and their mtdna are younger clades of L which have been founded just recently“Moreover, our hypothesis of an early return and subsequent expansion inside Africa of carriers of L3 and E haplogroups might help explain, the Neanderthal introgression detected in the western African Yoruba and in northern African Tunisian Berbers...”“black” Africans did not exist yet at that timeThe most frequent haplogroup in North Africa is E1b1b m81 also known as the Berber marker.
-
What is the step-by-step procedure to apply for an SBI PO and a clerk?
Here is how you can apply for SBI PO :As per SBI PO Eligibility, any Indian National who holds a Bachelor’s Degree can apply for SBI PO.All eligible candidates should visit the official website and register themselves via their mobile no. and email id.Next, upload scanned images of your photograph and signature in the prescribed format.Then, enter their personal and academic details to complete the form-filling process.Last, you must pay the requisite application fee. Candidates from General and OBC category have to payINR 600, while SC/ST/PWD candidates must pay INR 100.All candidates who register successfully can obtain SBI PO Admit Card from the official website.The Prelims rounds of SBI PO 2018 will be held in May 2018, while SBI PO Mains will be held in June. All selected candidates will proceed towards the GD/PI round that will be held in July 2018.How to Fill SBI PO Application Form 2018?SBI will release the application form in the month of April 2018. It must be noted that the entire registration process will be held online. There is no provision of obtaining SBI PO 2018 Application Form from offline mode.There are 5 broad steps you need to follow to fill SBI PO Application Form. They include:RegistrationUploading Photograph and SignatureApplication Form FillingFee PaymentConfirmationStep 1: RegistrationCandidates are required to follow the steps mentioned below to register themselves for SBI PO 2018Visit the official website i.e State Bank of IndiaClick for New registration option (written in yellow box)You can also click on the ‘How to Apply’ option provided in the Reference Documents to check the registration process.Disclaimer: Application form can be rejected if found ineligible at any point of time. The following message popup will be displayed for the information:Candidates must fulfil eligibility Criteria which is mentioned aboveOnline payment can only be made through: Debit Cards, Credit Cards, Internet Banking, IMPS, Cash cards and Mobile Wallets.The online registration process will only be completed after the submission of the application fees. Application fees can be submitted online as well as offline.If any Candidate wants to pay the SBI PO application fee through offline mode, then they can pay the application fee through bank challan. They can submit the fee by submitting SBI Challan.SBI (conducting Authority) can also allot any centre as per their convenience other than he/she has opted for.After reading all the information, candidates can press the continue buttonNext, SBI PO Registration Form will open. Enter the below-mentioned details:Name: Enter your full name properly (includes First name, Middle name and Last name). If candidates don’t have any Middle name then they can ignore that option. Name entered should be same as that of the given ID proof that you will show during the time of the examination as well as Interview.Mobile Number: Mobile number should remain the same till the entire procedure. No changes in the mobile number will be accepted later. No need to prefix 0.E- Mail ID: Enter your E- mail IDs correctly to get notifications regarding any update related to SBI PO examination or application procedure.After filling the basic information, candidates will be asked for the confirmation regarding the information provided by them and once you press the OK button, you cannot go back. So be careful while filling up the basic information.Press Ok to confirm your registration. You will be provided with registration number and password.Note down your registration number and password details to edit your application form (if necessary).Also to take a print of your submitted SBI PO application form simply login using registration number and password, and click on print/save button.Step 2: Upload Photograph and SignatureCandidates have to upload their recent passport size photograph and signature in SBI PO application form.Here are some instructions you need to adhere to:The photograph and signatures should be scanned properly and should be clearly visible.If there is some extra space around the scanned image that is unnecessary, it can be cropped to reduce the size.Both, the photograph and signature, have to be uploaded to the proper specifications.Signature must be done by using Black ink pen on a white sheet of paper.Photo and Signature Specifications for SBI PO 2018‘Choose File’ option will appear. Candidates can select their photograph and signature respectively by clicking at the options.Click at the next button and Window three will appear.Step 3: SBI PO Application Form FillingAfter uploading the images, enter your Qualification/ Academic details:Educational qualification should be filled correctly (as on 1.07.2018)The date of passing eligibility examination should be the date appearing on the mark sheet or provisional certificate issued by the University/Institute.Those who are in final year/ semester of their graduation may also apply provisionally subject to the condition that if they will be called for an interview, they must have cleared their Graduation before 1.07.2018Fractional percentage so arrived will be ignored i.e 59.99 will be treated as less than 60% and 54.99% will be treated as less than 55%.Fill the degree, name of the institution, place of the institution, date of passing, the percentage of marks and grade. Candidates have to choose the best possible option available.‘Other details option’ is also provided for the reserved classes if they are interested in participating for the Pre- exam training. This section is optional.After submitting all the details, preview option will appear where candidates can recheck the information provided by them and they can change/edit any incorrect information.Category and Code to be filled in SBI PO Application Form 2018Given below are the categories and codes which must be filled in the application form for SBI PO 2018. Note that candidates belonging to OBC (Non Creamy Layer) must categorize themselves in the General criteria only.Step 4: Application Fee PaymentAfter verifying all details and ensuring that they are accurate, candidate must proceed with the application fee payment.The fee can be paid using any of the online options, such as debit card, credit card or net banking.Fees for the General category and OBC is INR 600 and for SC/ST, it’s INR 100.Once the fee has been paid successfully, a receipt is generated.Candidate must save a copy of this receipt as proof of fee payment.For any further queries, Candidates can visit registration_helpdesk@ibpsorg.orgStep 5: ConfirmationAfter successful completion of payment method, you will get conformation message /mail by saying your application form acceptedCandidates are advised to take a printout of SBI PO 2018 Application Form for the future use.To take a print of your submitted SBI PO application form simply login using registration number and password, and click on print/save buttonThe copy of the completed form need not be sent to the bank.In case payment is unsuccessful:Candidate must register afresh and start the form filling process a second time.The Amount debited for failed transaction will be refunded back to your account within 3 -7 working days. The payment links are active 24 hours continuously. So if you still cannot make the fee payment during the day time, you can do it in the night also.Now that you know the process ups and downs you can start preparing for the same but how do we start for this. Now for these types of queries you can read below and find the best possible solutions for the same.You can get coaching from many institutes but it all depends on which coaching institute you are okay or comfortable to study and make sure that you understand everything and are able to see your growth in terms of knowledge and skills for clearing exams.Not just this but there are many other things that you must know before you start preparing through any specific ways.First let’s sort out how do you wish to prepare for the exam because that would only help you find the best institute or coaching for which you are looking. Thus I have elaborated following possible methods through which you can prepare in a way that will help you in clearing the exam and making the most out of it.There are many best ways now to pass this exam, well actually not just this exam but most of the exam because things are getting much easier and at the same time much competitive.THE BEST TIPEMOTION + INFORMATION= LONG-TERM MEMORYThis means if you have kind of emotion connected with any kind of information then it is the best thing possible because then it would become long-term memory which you will be able to remember in the exam too. The same reason you are able to remember a movie thus I would recommend for this part online video lectures links to which I am providing if you wish to learn.Eventually there will be no best way for preparing instead there will be many ways for preparing as of now because at the time when I was preparing there were not much ways for the exam preparation. All we had were offline classes to go and make the hectic schedule go on for the time which we were preparing. This had many downfall such as not able to make it up to the preparation over the week, not being able to make the study goals fulfilled because we had to spend time in the classes for studying the further concepts.Now here few things were to keep in mind that our previous preparation was left uncompleted because we had to study further for the new topics in the classes. The fear of not leaving the topics being taught we left the previous preparation in between at the time when I was preparing things were not much over the net.At the time when I was preparing things were not much over the net.We only had few options to prepare which was either going to tutions and taking classes or having a personal trainer which will teach you at home. But trust me guys no one had that much amount to give to a personal teacher even if it was someone who was financially well off from the family they also had to come in the classes.So we had to sacrifice our time of the job and work as part time employee over some where and then study along at the time when I was preparing things were not much over the net.Below for your comparison I will be mentioning all the ways through which you can practice studying but you can surely select the one that suits you the best.Now how do we start preparing for this,now for these things there are many other ways to prepare for the best to happen so in the best way possibly could be the one where you get the best quality of study but not just this there are many key factors as mentioned belowQuality Study MaterialTime to study on your ownBest Doubt session in which you definitely interact with the teachersBest ever test samplesAlso you must be able to make your scheduleNow all these things where do you think you can get. I will let you decide but also share my best way of preparation through which I prepare and made what I am today. Thus below I will share all the best possible ways to prepare.“A major part of your clearing exam depends upon the way you are preparing, as for the best results one most time should be utilized and least should be used anywhere”Thus according to the above information one should be able to make the decision the way they want to prepare.Now coming on to the main topic that through which way are you going to study for all this.Now here I am going to mention the following way to prepare which are as mentioned below-Offline Weekend Btach InstitutesOnline Sources such as YouTubeOnline Specialized InstituteHere I would also like to know through you which one you think is the best way to prepare for not just thins but any course or examination if you self determined to study on your own, which you eventually will have to do for the best results possible other wise the exam won’t be easy for you.Now according to a survey done on Facebook it was found that more than 80% of the students felt that Online Specialized Institutes were the best ways to prepare when were told with the pros and cons for each of them.Thus assuming the same over here but still I will be mentioning both the pros and cons for each of them so that you can also check by yourself which one suits your life better and which one not.If one goes for the first option then he/she will have to bear the following things as mentioned below -High fee amountSimilar content as othersLess duration of the coursesTravelling expensesNo self-planning strategyAlong with all these problems, one might have to face the problem of not having complete content as these institutes might not complete the content properly because of being in hurry of completing the course in the duration mentioned.But if one opts for free sources online they will find mostly the same content over the internet but there is no confirmation that the content will be correct or not also saying that if the content is there is not sure that it will be in structured form and it will be providing proper guidance step wise but there is nothing problem in that as one always have another option .Now if you go for the last and best option of Online Coaching Institute then below mentioned are the things one will have to bear -Less fee amountNo traveling expensesBooks/E-booksOne year validity of courseCertificate ( after the examination)Active doubt sessionChat support with facultyCall supportPractical sessionsNow while I had no chance for clearing my exam I came across this institute mentioned below and here I am cleared by 12th with 70% not bragging about this, but at that time even teachers rejected from taking me into their batches maybe because I was late for the batches now.Takshila Learning Pvt LtdWebsite- www.takshilalearning .comAnd do fill the form for best assistance and knowledge along the website.This is what I did for my preparation in the best possible ways but you can also prepare for yourself in your own way which you feel is the best.Thanks
-
What are probabilistic graphical models, and why are they useful?
These are Probabilistic Graphical Models. They are arguably our most complete and promising toolkit for inferring truth from complexity. They're born from a single set of principles that endow our machines to dominate chess, diagnose disease, translate language, decipher sound, recognize images and drive cars. 'Neural Networks' and 'Probabilistic Programming' are famous signatures of the Machine Learning community simply because they are effective tool sets for applying these devices.My aim here is to reveal the machinery behind this magic. I intend to show what they are, why we use them and how we actually use them. To do that, I’ve answered seven questions on this subject:What are Probabilistic Graphical Models and why are they useful?What is 'exact inference' in the context of Probabilistic Graphical Models?What is Variance Inference in the context of Probabilistic Graphical Models?How are Monte Carlo methods used to perform inference in Probabilistic Graphical Models?How are the parameters of a Bayesian Network learned?How are the parameters of a Markov Network learned?How is the graph structure of Probabilistic Graphical Models learned?I realize this is a ton to digest, especially for internet browsing, but allow me to sell you. This information is typically delivered with a worthwhile 1000+ page textbook to graduate computer scientists. We can 80/20 these ideas with just a few answers! It'll take discipline, but you'll gain a surprisingly good understanding of an absolutely foundational theory of Machine Learning.As a compromise, I've structured things such that you need only read a subset of these answers to get a full picture. Here's a map of that structure:For example, if you read [math]1 \rightarrow 2 \rightarrow 6 \rightarrow 7[/math], you'll get a complete taste. Also, I'll include refreshers at the beginning of each answer - this should make things more self contained. (If you read these answers in sequence, I'd skip those refreshers, as they will sound redundant.)If this sounds like a good deal to you, please follow those questions!Now, let's start walking.Notation GuideAs a first stop, we'll review notation, an admittedly boring place. But, it's my unconventional belief that most confusion is due to notation. So if we wish to survive, we'll need a few tips:An upper case non-bold letter indicates a single random variable ('RV'). The same letter lower cased with a super script indicates a specific value that RV may take. For example, [math]X=x^1[/math] is the event the RV [math]X[/math] took on the value [math]x^1[/math]. We call this event an assignment. The set of unique values an RV may take is [math]Val(X)[/math]. So we might have [math]Val(X)=\{x^0,x^1\}[/math] in this case.A bold upper case letter indicates a set of RVs (like [math]\mathbf{X}[/math]) and a bold lower case letter indicates a set of values they may take. For example, we may have [math]\mathbf{X}=\{A,B\}[/math] and [math]\mathbf{x}=\{a^3,b^1\}[/math]. Then the event [math]\mathbf{X}=\mathbf{x}[/math] is the event that [math]A=a^3[/math] happens and [math]B=b^1[/math] happens. Naturally, [math]Val(\mathbf{X})[/math] is the set of all possible unique joint assignments to the RVs in [math]\mathbf{X}[/math].If you see [math]\mathbf{x}[/math] (or [math]\mathbf{y}[/math] or [math]\mathbf{z}[/math] etc...) within a probability expression, like [math]P(\mathbf{x}|\cdots)[/math] or [math]P(\cdots|\mathbf{x})[/math], that's always an abbreviation of the event '[math]\mathbf{X}=\mathbf{x}[/math]'.Perhaps confusingly, we also abbreviate the event '[math]\mathbf{X}=\mathbf{x}[/math]' as '[math]\mathbf{X}[/math]', though this isn't a clean abbreviation. Omission of [math]\mathbf{x}[/math] means one of two things: either we mean this for any given [math]\mathbf{x}[/math] or for all possible [math]\mathbf{x}[/math]'s. As an example for the latter case, 'calculate [math]P(\mathbf{X})[/math]' would mean calculate the set of probabilities [math]P(\mathbf{X}=\mathbf{x})[/math] for all [math]\mathbf{x}\in Val(\mathbf{X})[/math].[math]\sum_\mathbf{X}f(\mathbf{X})[/math] is shorthand for [math]\sum_{\mathbf{x}\in Val(\mathbf{X})}f(\mathbf{X}=\mathbf{x})[/math]. This is similarly true for [math]\prod_\mathbf{X}(\cdot)[/math] and [math]\textrm{argmin}_\mathbf{X}(\cdot)[/math]. Look out for this one - it can sneak in there and change things considerably.You may see equations like [math]f(A,B,C)=g(\mathbf{X})h(\mathbf{Y})[/math]. They look strange - the RVs on the left aren't on the right! Well, in such cases, you also have something like [math]\mathbf{X} = \{A,B\}[/math] and [math]\mathbf{Y} = \{B,C\}[/math]. So the equation really is [math]f(A,B,C)=g(A,B)h(B,C)[/math].Probability distributions are references with a [math]P[/math], [math]\textrm{Q}[/math], [math]q[/math] or [math]\pi[/math]. Keep in mind that distributions are a special kind of function. Remember that!Everything is in reference to the discrete case. Unfortunately, the continuous case is not a simple generalization from the discrete case. The minor exception is in the visuals. The discrete case is less friendly to graphs, so I might use some continuous distributions. As it relates to the discussion, pretend these are in fact discrete distributions with a fine granularity.Almost all of this notation comes from the text Probabilistic Graphical Models - one of those 1000 page monsters. That book is extremely thorough, and should be considered stop number 8.Still here? You must have discipline! Onto the fun stuff - we ask:What generic problem do PGMs address?Our goal is to understand a complex system. We assume the complex system manifests as [math]n[/math] RVs, which we may write as [math]\mathcal{X} = \{X_1,X_2,\cdots,X_n\}[/math] [1][2]. We take it that 'a good understanding' means we can answer two types of questions accurately and efficiently for these RVs. If we say [math]\mathbf{Y}[/math] and [math]\mathbf{E}[/math] are two given subsets of [math]\mathcal{X}[/math], then those questions are:Probability Queries: Compute the probabilities [math]P(\mathbf{Y}|\mathbf{E}=\mathbf{e})[/math]. That is, what is the distribution of the RV's of [math]\mathbf{Y}[/math] given we have some observation of the RVs of [math]\mathbf{E}[/math]?MAP Queries: Determine [math]\textrm{argmax}_\mathbf{Y}P(\mathbf{Y}|\mathbf{E}=\mathbf{e})[/math]. That is, determine the most likely assignments of RVs given an assignment of other RVs.Before continuing, we should point a few things out:Since [math]\mathbf{Y}[/math] and [math]\mathbf{E}[/math] are any two subsets of [math]\mathcal{X}[/math], there is potentially a remaining set (call it [math]\mathbf{Z}[/math]) that's in [math]\mathcal{X}[/math]. In other words, [math]\mathbf{Z} = \mathcal{X} - \{\mathbf{Y},\mathbf{E}\}[/math]. This set appears left out of our questions, but is very much at play. We have to sum these RVs out, which can considerably complicate our calculations. For example, [math]P(\mathbf{y}|\mathbf{e})[/math] is actually [math]\sum_\mathbf{Z}P(\mathbf{y},\mathbf{Z}|\mathbf{e})[/math].We haven't mentioned any model yet. This set up is asking generically for probabilities and values that accurately track reality.To this end, we are assisted by the fact that we have some, at least partial, joint observations of [math]\mathcal{X}[/math]. However, some of our [math]n[/math] RVs may never be observed. These are called 'hidden' variables and they will complicate our lives later on.This set up is extremely general, and as such, this problem is extremely hard.The problem with joint distributions.Our starting point, perhaps surprisingly, will be to consider the joint distribution of our RVs [math]\mathcal{X}[/math], which we aren't given in real application (but we'll get there). We'll call that joint distribution [math]P[/math]. Conceptually, we can think of this as a table that lists out all possible joint assignments of [math]\mathcal{X}[/math] and their associated probabilities. So if [math]\mathcal{X}[/math] is made up of 10 RVs, each of which can take 1 of 100 values, this table has [math]100^{10}[/math] rows, each indicating a particular assignment of [math]\mathcal{X}[/math] and it's probability.The issue is, for a complex system, this table is too big. Even if we had the crystal ball luxury of having [math]P[/math], we can't handle it. So now what?The Conditional Independence statementWe need a compact representation of [math]P[/math] - something that gives us all the information of that table, but doesn’t involve writing it down. To this end, our saving grace is the Conditional Independence (CI) statement:Given subsets of RVs [math]\mathbf{X}[/math], [math]\mathbf{Y}[/math] and [math]\mathbf{Z}[/math] from [math]\mathcal{X}[/math], we say [math]\mathbf{X}[/math] is conditionally independent of [math]\mathbf{Y}[/math] given [math]\mathbf{Z}[/math] if[math]P(\mathbf{x},\mathbf{y}|\mathbf{z})=P(\mathbf{x}|\mathbf{z})P(\mathbf{y}|\mathbf{z})\tag*{}[/math]for all [math]\mathbf{x}\in Val(\mathbf{X})[/math], [math]\mathbf{y}\in Val(\mathbf{Y})[/math] and [math]\mathbf{z}\in Val(\mathbf{Z})[/math]. This is stated as '[math]P[/math] satisfies [math](\mathbf{X}\perp \mathbf{Y}|\mathbf{Z})[/math][3]'Now, if we had sufficient calculation abilities, we could calculate the left side and the right side for a distribution [math]P[/math]. If the equations hold for all values, then, by definition, the CI statement holds. Intuitively, though not obviously, this means that if you are given the assignment of [math]\mathbf{Z}[/math], then knowing the assignment of [math]\mathbf{X}[/math] will never help you guess [math]\mathbf{Y}[/math] (and vice versa). In other words, [math]\mathbf{X}[/math] provides no information for predicting [math]\mathbf{Y}[/math] beyond what [math]\mathbf{Z}[/math] has. Similarly, you can't predict [math]\mathbf{X}[/math] from [math]\mathbf{Y}[/math] any better.Knowing such statements turns out to be massively useful - they give us that compact representation we need. To see this, let's say [math](X_i \perp X_j)[/math] for all [math]i \in \{1,\cdots,10\}[/math] and [math]j \in \{1,\cdots,10\}[/math] where [math]i\neq j[/math]. This is to say, all RVs are independent of all other RVs. It turns out that with these statements, we only need to know the marginal probabilities of each value for each RV (which is a total of [math]10\cdot 100=1000[/math] values) and may reproduce all the probabilities of [math]P[/math]. So if we are considering the case where [math]\mathbf{X}=\mathcal{X}[/math] and would like to know the probability [math]P(\mathbf{X}=\mathbf{x})[/math], we simply return [math]\prod_{i=1}^{10}P(X_i=x_i)[/math], where [math]x_i[/math] is the [math]i[/math]-th element of [math]\mathbf{x}[/math].Though this isn't just a save on storage. This is a simplification on [math]P[/math] that will ease virtually any interaction with [math]P[/math], including summing over many assignments and finding the most likely assignment. So at this point, I'd like you to think that CI statements regarding [math]P[/math] are a requirement for wielding it.Now put a pin in this and let's switch gears.The Bayesian NetworkIt's time to introduce the first type of PGM - the Bayesian Network ('BN'). A BN refers to two things, both in relation to some [math]\mathcal{X}[/math]: a BN graph (called [math]\mathcal{G}[/math]) and an associated probability distribution [math]P_B[/math]. [math]\mathcal{G}[/math] is a set of nodes, one for each RV of [math]\mathcal{X}[/math], and a set of directed edges, such that there are no directed cycles. Said differently, it's a DAG. [math]P_B[/math] is a distribution with probabilities for assignments of [math]\mathcal{X}[/math] using a certain rule and Conditional Probability Tables ('CPTs' and 'CPDs'), which augment [math]\mathcal{G}[/math]. That rule, called the 'Chain Rule for BNs', for determining probabilities can be written:[math]P_B(X_1,\cdots,X_n)=\prod_{i=1}^n P_B(X_i|\textrm{Pa}_{X_i}^\mathcal{G})\tag*{}[/math]where [math]\textrm{Pa}_{X_i}^\mathcal{G}[/math] indicates the set of parent nodes/RVs of [math]X_i[/math] according to [math]\mathcal{G}[/math]. The CPDs tell us what the [math]P_B(X_i|\textrm{Pa}_{X_i}^\mathcal{G})[/math] probabilities are. That is, a CPD lists out the probabilities of all assignments of [math]X_i[/math] given any joint assignment of [math]\textrm{Pa}_{X_i}^\mathcal{G}[/math][4]. These CPDs are the parameters of our model. Their form is to list out actual conditional probabilities from [math]P_B[/math].To help, let's consider a well utilized example from that monstrous text: the 'Student Bayesian Network'. Here, we're concerned with a system of five RVs: a student's intelligence ([math]I[/math]), their class's difficulty ([math]D[/math]), their grade in that class ([math]G[/math]), their letter of recommendation ([math]L[/math]) and their SAT score ([math]S[/math]). So [math]\mathcal{X}=\{I,D,G,L,S\}[/math]. The BN graph along with the CPDs can be represented as:According to our rule, we have that any joint assignment of [math]\mathcal{X}[/math] factors as:So we would calculate a given assignment as:Not too bad, right? All this is to show is that a BN along with CPDs gives us a way to calculate probabilities for assignments of [math]\mathcal{X}[/math].Now we're ready for:The big idea.It's so big, it gets its own quote block:The BN graph, just those nodes and edges, implies a set of CI statements regarding it's accompanying [math]P_B[/math].It's a consequence of the Chain Rule for calculating probabilities. As a not-at-all-obvious result, a BN graph represents all [math]P[/math]'s that satisfy these CI statements and each of those [math]P[/math]'s could be attained with an appropriate choice of CPDs.For a BN, one form of those CI statements are:[math](X_i \perp\textrm{NonDescendants}_{X_i}|\textrm{Pa}_{X_i}^\mathcal{G})[/math] for [math]X_i \in \mathcal{X}[/math]So in the student example, we'd have this set:The third statement tells us that if you already know the student's intelligence and their class's difficulty, then knowing their SAT score won't help you guess their grade. This is because the SAT score is correlated with their grade only via their intelligence, and you already know that.These are referred to as the local semantics of the BN graph. To complicate matters, there are almost always many other true CI statements associated with a BN graph outside of the local semantics. To determine those by inspecting the graph, we use a scary 'D-separation' algorithm that I will shamelessly not explain.There is a reason this is so important. Since a BN graph is a way of representing CI statements and such statements are a requirement for handling a complex system's joint distribution (if you had it), then this is a good reason to use a BN to represent such systems. If we can accurately represent a system with a BN, we will be able to calculate our probability and MAP queries. Therefore, BNs will solve our problems when we're dealing with a certain class of [math]P[/math]'s. This choice, unsurprisingly, is called our representation.But there's an issue - I said a 'class' of [math]P[/math]'s. It's not hard to invent [math]P[/math]'s that come with CI statements a BN cannot represent.So now what? Well, we have other tools, the biggest of which is...The Markov NetworkA Markov Network ('MN') is likewise composed of a graph ([math]\mathcal{H}[/math]) and a probability distribution ([math]P_M[/math]). Though this time, the graph's edges are undirected and it may have cycles. The consequence is that a MN can represent a different set of CI statements. But, the lack of directionality means we can no longer use CPDs. Instead, that information is delivered with a factor, which is a function (function! remember it) that maps from an assignment of some subset of [math]\mathcal{X}[/math] to some nonnegative number. These factors are used to calculate probabilities with the 'Gibbs Rule'[5].To understand the Gibbs Rule, we must define a complete subgraph. A ‘subgraph’ is exactly what it sounds like - we make a subgraph by picking a set of nodes from [math]\mathcal{H}[/math] and including all edges from [math]\mathcal{H}[/math] that are between nodes from this set. A 'complete' graph is one which has every edge it can - each node has an edge to every other node.Now, let's say [math]\mathcal{H}[/math] breaks up into a set of [math]m[/math] complete subgraphs. By 'break up', I mean that the union of all nodes and edges across these subgraphs gives us all the nodes and edges from [math]\mathcal{H}[/math]. Let's write the RVs associated with the nodes of these subgraphs as [math]\{\mathbf{D}_i\}_{i=1}^m[/math]. Let's also say we have one factor (call it [math]\phi_i(\cdot)[/math]) for each of these. We refer to these factors together with [math]\Phi[/math], so [math]\Phi=\{\phi_i(\cdot)\}_{i=1}^m[/math]. For terminology's sake, we say that the 'scope' of the factor [math]\phi_i(\cdot)[/math] is [math]\mathbf{D}_i[/math] because [math]\phi_i(\cdot)[/math] takes an assignment of [math]\mathbf{D}_i[/math] as input.Finally, the Gibbs Rule says we calculate a probability as:where(It's hidden from this notation, but we're assuming it's clear how to match up the assignment of [math]X_1,\cdots,X_n[/math] with the assignments of the [math]\mathbf{D}_i[/math]'s.)Wait - the MN was introduced because it represents a different set of CI statement. So, which ones? It's considerably simpler in the case of a MN. A MN implies the CI statement [math](\mathbf{X} \perp \mathbf{Y}|\mathbf{Z})[/math] if all paths between [math]\mathbf{X}[/math] and [math]\mathbf{Y}[/math] go through [math]\mathbf{Z}[/math]. Easy!Now let's get specific. Below is an MN for the system [math]\mathcal{X}=\{A,B,C,D\}[/math] and the CI statements it represents:As you may notice, it's not hard to write those CI statements by viewing the graph.While we're here, let's write out the Gibbs Rule. By looking at this, we could identify our complete subgraphs as: [math]\{\{A,B\},\{B,C\},\{C,D\},\{D,A\}\}[/math]. With that, we calculate a probability as:whereTo repeat, each [math]\phi_i(\cdot,\cdot)[/math] is just a function that maps from it's given joint assignment to some nonnegative. So if [math]A[/math] and [math]B[/math] could only take on two values each, [math]\phi_1(\cdot,\cdot)[/math] would relate the four possible assignments to four nonnegative numbers. These functions serve as our parameters just as the CPDs did. Determining these functions brings us from a class of [math]P[/math]'s to a specific [math]P[/math] within it, defined with probabilities.But, ahem, uhh… there's an issue. In the BN case, I said:As a not-at-all-obvious result, a BN graph represents all [math]P[/math]'s that satisfy its CI statements and each of those [math]P[/math]'s could be attained with an appropriate choice of CPDs.The analogous is not true in the case of MNs. There may exist a [math]P[/math] that satisfies the CI statements of a MN graph, but we can't calculate it's probabilities with the Gibbs rule. Damn!Fortunately, these squirrely [math]P[/math]'s falls into a simple, though large, category: those which assign a zero probability to at least one assignment. This leads us to the Hammersley-Clifford theorem:If [math]P[/math] is a positive distribution ([math]P(\mathbf{X}=\mathbf{x})>0[/math] for all [math]\mathbf{x} \in Val(\mathcal{X})[/math]) which satisfies the CI statements of [math]\mathcal{H}[/math], then we may use the Gibbs Rule, along with a choice of complete subgraphs and associated factors, to yield the probabilities of [math]P[/math]. [6]And that about does it for the basics of MNs. They are just another way of representing another class of [math]P[/math]'s.How do BNs and MNs compare?At this point, we're not evolved enough for a full comparison, so let's do a partial one.First, it's clearly easier to determine CI statements in a MN - no fancy D-separation algorithm required. This follows from their simple symmetric undirected edges, which make them a natural candidate for certain problems. Broadly, MNs do better when we have decidedly associative observations - like pixels on a screen or concurrent sounds. BNs are better suited when we suspect the RVs attests to distinct components of some causal structure. Timestamps and an outside expectation of what's producing the data are helpful for that.Also, there's a certain overlap between a MN and a BN that'll unify our discussion in later answers. That is, the probabilities produced by the Chain Rule of any given BN can be exactly reproduced by the Gibbs Rule of a specially defined MN. To see this, look at the Chain Rule - [math]P_B(X_i|\textrm{Pa}_{X_i}^\mathcal{G})[/math] is just the conditional probability of some (unspecified) [math]X_i[/math] value given some assignment of the parent RVs. Well, to translate this to the Gibbs Rule, let [math]\mathbf{D}_i=\{X_i\}\cup\textrm{Pa}_{X_i}^\mathcal{G}[/math]. Next, define [math]\phi_i(\mathbf{D}_i)[/math] to produce the same output you'd get from looking up the BN conditional probability in the CPD (which is [math]P_B(X_i|\textrm{Pa}_{X_i}^\mathcal{G})[/math]). Awesome - now the Gibbs Rule is the same expression as the Chain Rule. This is useful because we can speak solely in terms of the Gibbs Rule and whatever we discover, we know will also work for the Chain Rule (and hence BNs). What this doesn't mean is that MNs are a substitute for BNs. If you were to look at this invented MN, it would likely imply way more edges in its graph and therefore, fewer CI statements and therefore, a wider and more unwieldy class of [math]P[/math]'s. In other words, BNs are still useful representations.But there's more to learn.Let's say we determined our graphical model along with its parameters. How do we actually answer those queries? Well, I have three suggestions:2. What is 'exact inference' in the context of Probabilistic Graphical Models?3. What is Variance Inference in the context of Probabilistic Graphical Models?4. How are Monte Carlo methods used to perform inference in Probabilistic Graphical Models?Footnotes[1] This is the one exception where we don't refer to a set of RVs with a bold uppercase letter.[2] This actually isn't the fully general problem specification. In complete generality, the set of RVs should be allowed to grow/shrink over time. That, however, is outside what I expect to accomplish in these posts.[3] There is a subtlety of language here. Often we'll say '[math]P[/math] satisfies these CI statements'. That means those CI statements are true for [math]P[/math], but others may be true as well. So it means 'these CI statements' are a subset of all [math]P[/math]'s true CI statements. This technicality matters, so keep an eye out for it.[4] If [math]X_i[/math] doesn't have any parents, then the CPD is the unconditional probability distribution of [math]X_i[/math].[5] This isn't a real name I'm aware of, but the form of that distribution makes it a Gibbs distribution and I'd like to maintain an analogy to BNs, which had the Chain Rule.[6] The implication goes the other way as well: If the probabilities of [math]P[/math] can be calculated with the Gibbs Rule, then it's a positive distribution which satisfies CI statements implied by a graph which has complete subgraphs of RVs that correspond to the RVs of each factor. This direction, however, doesn't fit into the story I'm telling, so it sits as a lonely footnote.Sources[1] Koller, Daphne; Friedman, Nir. Probabilistic Graphical Models: Principles and Techniques (Adaptive Computation and Machine Learning series). The MIT Press. Kindle Edition. This is the source of the notation, the graphics in this answers (with permission) and my appreciation for this subject.
Trusted esignature solution— what our customers are saying







Get legally-binding signatures now!
Frequently asked questions
How do i add an electronic signature to a word document?
How do you sign financial documents in pdf?
airSlate SignNow how to sign a pdf?
Get more for Edit eSignature Form Later
- How Can I Electronic signature Virginia Plumbing Document
- How Do I Electronic signature Virginia Plumbing Word
- How To Electronic signature Virginia Plumbing Document
- How Do I Electronic signature Virginia Plumbing Document
- Can I Electronic signature Virginia Plumbing Document
- Help Me With Electronic signature Virginia Plumbing Word
- Help Me With Electronic signature Virginia Plumbing Document
- How Can I Electronic signature Virginia Plumbing Document
Find out other Edit eSignature Form Later
- Sample letter promotional 497333566 form
- Master grower contract templates form
- Sample letter for high school fundraiser form
- Sole proprietorship 497333569 form
- Letter fees form
- Letter thanking form
- Model letter accountants to auditors form
- Software distribution agreement form
- Sample letter family 497333574 form
- Electronic agreement form
- Sample letter party form
- Agreement teacher contract form
- Tax receipt form
- Agreement parents form
- Sample letter for request for donated items form
- Irrevocable trust death form
- Jury duty letter form
- Duty report letter in english form
- Irrevocable trust children form
- Request during form